Conodonta
Pour l'homonyme †Conodontes Laugel, 1862, voir †Trogontherium Fischer von Waldheim, 1809, un genre de castors préhistoriques.
Ne pas confondre avec Eutriconodonta, un taxon de mammifères préhistoriques.

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Super-classe | Agnatha (polyphylétique) |
| Clade | Cyclostomata ? |
Taxons de rang inférieur
Le terme Conodonta référe à des formes denticulées fossiles apparues au début du Cambrien, il y a 529 Ma, et disparaissant immédiatement après la fin du Trias, il y a 200 Ma, lors d'une extinction commune à cette fin de période. Les animaux vivants devaient ressembler à des anguilles primitives, mais sans mâchoires.
Francisé en conodontes, le terme en latin scientifique vient du grec ancien kōnos signifiant « cône » et ὀδούς ou odoús signifiant « dent », soit « dents en cônes »).
Perspective historique et description

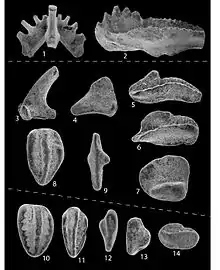
Fig. 1. Acontiodus latus, b von den Seitenflächen, c im Profil von der concaven Seite.
Fig. 2. Acontiodus gracilis, a von der concaven Seite, b und c die Seitenflächen.
Fig. 3. Scolopodus sublaevis, a und b die beiden Seitenflächen.
Fig. 4. Scolopodus semicostatus, a und b die beiden Seitenflächen.
Fig. 5. Scolopodus aequilateralis, a von dem concaven, b von dem concaven Rande angesehn, b die eine Seitenfiäche.
Fig. 6. Scolopodus quadratus, b und c die beiden Seitenflächen, a im Profil.
Fig. 7. Scolopodus costatus, a vom concaven Rande, b und c, die beiden Seitenflächen.
Fig. 8. Scolopodus striatus, a vom concaven d vom concaven Rande, b und c die beiden Seitenflächen.
Fig. 9. Drepanodus acutus, eine Seitenfläche.
Fig. 10. Machairodus rhomboideus, eine Seitenfläche.
Fig. 11. Drepanodus obtusus, eine Seitenfläche.
Fig. 12. Machairodus rhomboideus var ., eine Seitenfläche.
Fig. 13. Acodus crassus, a und b die beiden Seitenflächen.
Fig. 14. Machairodus dilatatus, von einen Seite.
Fig. 15. Machairodus solidus, eine Seitenfläche.
Fig. 16. Drepanodus inflexus, eine Seitenfläche.
Fig. 17, Fig. 18 und Fig. 19. Oistodus lanceolatus, Fig. 17 eine Seitenfläche. Fig. 18 a im Profil von dem concaven Rande angesehen, b und c die beiden Seitenflächen, Fig. 19 b und c die beiden Seitenflächen, c die Basis von unten betrachtet.
Fig. 20. Oistodus acuminatus, a und 6 die beiden Seitenflächen.
Fig. 21. Belodus gracilis, von einer Seite.
Fig. 22 und 23. Prioniodus elegans, a und b die beiden Seitenflächen.
Fig. 24. Prioniodus sulcatus, a und 8 die beiden Seitenflächen.
Fig. 25. Prioniodus carinatus, eine Seitenfläche.
Fig. 26 bis Fig. 31. Cordylodus angulatus, verschiedene Varietäten.
Fig. 32-33. Cordylodus rotundatus, a die beiden Seitenflächen, b die Basis.
Fig. 34. Cordylodus angulatus.
Fig. 35. Acontiodus triangulosis, a der concave Rand, 6 und c die Seitenflächen, d Ansicht der Basis von unten.
Fig. 36. Machairodus ensiformis, a und 6 die beiden Seitenflächen.
Fig. 37. Oistodus inaequalis, a und b die beiden Seitenflächen.
Fig. 38. Machairodus inaequalis, a und b die beiden Seitenflächen, c die Basis von unten angesehen.
Fig. 39. Machairodus planus, eine Seitenfläche.
Fig. 40. Oistodus parallelus, eine Seitenfläche.
Initialement, ils étaient connus seulement à travers des microfossiles de dents. L'utilisation de ces "dents" devaient probablement servir d'appareils d'alimentation par filtration ou de système pour attraper et écraser.
La nature exacte de ces énigmatiques fossiles, jusque-là considérés comme des « vers », faisait alors l'objet de multiples controverses, notamment concernant l'absence de ce que Richard Owen appelle les "tubes calcigères", décrits dans les dents de vertébrés dans son livre Odontography écrit de 1840 à 1845.
En 1856, dans sa monographie sur les poissons fossiles[1], Christian H. Pander reconnaît dans certains « vers fossiles » du Trias un type de chordés particulier, le groupe des conodontes. Dans les termes employés par Pander lui-même, il s'agit de la dentition (structure maxillaire) de « poissons ». Il décrit des "dents" où on peut distinguer une pointe solide et une base creuse formant une "cavitas pulpae"[2]. Pander cite 14 noms de genres et des espèces associées.
En 1930, Wilhelm Eichenberg, géologue allemand, propose le nom de Conodontophorida, au rang de l'ordre, pour prendre en compte l'aspect de l'animal dans son intégralité (le sujet porteur de conodontes) et pas seulement celui de ses "dents"[3]. Mais, selon certains auteurs, ce terme est synonyme de Conodonta. Au cours des années 1930, le concept d'assemblage de conodontes a été décrit par Hermann Schmidt[4] et par Harold W. Scott[5], puis par d'autres auteurs dans les décennies qui ont suivi[6],[7],[8],[9].
Dans les années 1940, alors que le géologue et paléontologue américain Wilbert H. Hass s'attache à décrire la morphologie ultrastructurale[2], la composition chimique et l'orientation des cristaux des conodontes[10], d'autres poursuivent les travaux d'identification des espèces[11],[12],[13].

Dans les années 1980, la position cladistique des conodontes était inconnue jusqu'à ce que des conodontes soient retrouvés dans un Lagerstätte à Granton, près d'Édimbourg en Écosse, dans une strate du Carbonifère, associés à des tissus fossilisés[14],[15]. Les fossiles présentaient un appareil complet des pièces de conodontes au niveau de ce qui serait leur tête. Ces vertébrés auraient mesuré dans la quarantaine de millimètres en moyenne pour des conodontes de très petite taille (0,1 à 2 mm). Ils auraient possédé des yeux et auraient ressemblé aux anguilles actuelles (en bien plus petit). Ils pouvaient mesurer jusqu'à 40 cm pour les plus grands individus (cas de Promissum). Leurs grands yeux avaient une position latérale de ce qui rend peu probable un rôle prédateur. La musculature conservée laisse entendre que certains conodontes (Promissum au moins) étaient efficaces, mais incapables d'atteindre des pointes de vitesse.
En 1963, Lars E. Fåhraeus, un paléontologue suédois, publie un article sur la priorité nomenclaturale, restituant à Christian Heinrich von Pander (1794-1865), la paternité du nom des conodontes dans sa monographie des poissons fossiles parue en 1856[16].
Au début du XXIe siècle, 550 noms de genres valides sont connus, mais la réinterprétation en assemblages multi-éléments peut faire baisser ce nombre[17].
Dans les années 2010, les chercheurs classent définitivement les conodontes dans l'embranchement des chordés en se basant sur la présence d'ailerons, de rayons au niveau des nageoires, de muscles en forme de chevrons et d'une chorde.
Certains chercheurs les considèrent comme des vertébrés similaires en apparence aux myxines et lamproies modernes (groupe des cyclostomes), bien que l'analyse phylogénétique suggère qu'ils sont très éloignés de ces groupes. L'application de techniques récentes de spectrométrie (telles que la spectroscopie de rayons X à dispersion d'énergie ou la spectrométrie photoélectronique X) permettant de détecter la présence de kératine à l'état fossile, sous la forme de résidus soufrés, conduit à l'inverse à les mettre à la base du groupe des cyclostomes[18].
« Dents », « éléments » et « appareils »

_of_the_Illinois_basin_(1958)_(20654535006).jpg.webp)
Si l'on parle de dents dans la description de leur anatomie, celles-ci n'étaient pas portées par des mâchoires, comme chez les vertébrés gnathostomes actuels, mais étaient localisées au niveau des tissus mous de la bouche et du pharynx. Certains auteurs préfèrent donc utiliser le terme d'éléments pour les désigner.
Les éléments de conodontes sont formés de pointes recouvertes par un émail et d'une base en dentine[19].
Les différentes formes de conodontes sont décrites comme de simple cônes (ressemblant à des dents aigues), des barres (portant des structures en forme de pointe le long de leur bord), des lames (des rangées de cônes aplaties) ou des plateformes (comme des lames, avec des bords élargis des deux côtés).
Les conodontes sont trouvés seuls ou regroupés en "appareil", de manière souvent symétrique si l'appareil est bien conservé. La systématique est actuellement en train de subir des évolutions essayant de regrouper les genres et espèces en appareils de conodontes et en non-conodontes proprement dits.
Conodontes multi-éléments

Les appareils des conodontes peuvent comporter un certain nombre d'éléments. Selon leur position et leur forme, ceux-ci portent les noms de spathognathiforme, ozarkodiniforme, trichonodelliforme, néoprioniodiforme, et autres noms[20].
Éléments des ozarkodinides
Les appareils des ozarkodinides sont composés sur le devant d'un élément axial Sa, flanqué par deux groupes de quatre éléments allongés Sb et Sc qui devaient être inclinés obliquement. Au-dessus de ces éléments se trouve une paire d'éléments M (makellés) arqués et pointant vers l'intérieur. À l'arrière, les éléments S-M sont positionnés transversalement et s'opposent bilatéralement aux éléments pectiniformes (c.à.d. en forme de peigne) Pb et Pa[21].
Composition chimique
Les conodontes fossilisent généralement en francolite (mélange carbonate-apatite fluoré)[10] répondant à la formule suivante :
Ca5 Na0.14 (PO4)3.01 (CO3)0.16 F0.73 (H2O)0.85[22],[23],[24]
Index d'altération des conodontes

L'index d'altération des conodontes ("Conodonte Alteration Index" en anglais ou CAI) est un outil pour estimer le maximum de température atteint par une roche sédimentaire qui utilise l'altération thermique des fossiles de conodontes. Les conodontes présents dans les roches carbonatées fossilifères sont préparés par dissolution de la matrice avec un acide faible, puisque les conodontes sont composés d'apatite et ne se dissolvent pas aussi vite que les carbonates. Les fossiles sont alors comparés à l'index sous observation au microscope. Cet index a été développé par Anita Epstein et ses collègues de l'Institut d'études géologiques des États-Unis (United States Geological Survey ou USGS)[25].
L'index va de 1 à 6, comme suit:
| CAI | Couleur approximative du conodonte | Gamme de température (°Celsius) | |
|---|---|---|---|
| 1 | brun pâle | <50°-80° | |
| 2 | brun foncé | 60°-140° | |
| 3 | gris-brun foncé | 110°-200° | |
| 4 | gris foncé | 190°-300° | |
| 5 | noir | 300°-480° | |
| 6 | gris pâle à blanc | 360°-550° | |
L'index est utilisé communément par les paléontologues du fait de sa facilité de mesure et du fait de l'abondance des conodontes dans les dépôts de carbonates marins du Paléozoïque. Cependant, ces organismes ont disparu après le Trias et l'index n'est donc pas disponible en tant que technique pour analyser des roches plus récentes que 200 millions d'années. L'index peut être faussé positivement dans des régions d'altération hydrothermale[25],[26].
Histoire géologique
Le genre de protoconodontes Protohertzina est connu d'avant le Tommotien (c'est-à-dire de l'extrême fin du Précambrien)[27],[28].
Les euconodontes font leur apparition au Cambrien moyen.
Extinction
La disparition finale des conodontes s'est produite de façon échelonnée dans le temps en fonction des zones paléogéographiques considérés.
Ainsi, s'il est possible d'approximer de façon générale leur disparition avec la fin du Trias et donc le début du Jurassique, dans le détail, cette extinction est diachrone.
En effet, cette extinction se produit plus rapidement par exemple dans les zones côtières de la Néotéthys, durant la fin du Rhétien, que dans les océans profonds de la Panthalassa (par exemple: à Inuyama, au Japon) ou dans les bassins sédimentaires les plus isolés (par exemple: à Csővár, en Hongrie).
Dans ces deux derniers types d'environnement, l'extinction se produit donc plus tard, c'est-à-dire au Jurassique Inférieur pendant la première partie de l'Hettangien.
Les deux dernières espèces identifiées de conodontes sont Misikella posthernsteini et Neohindeodella detrei.
Systématique et phylogénie
| Craniata |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
- Notes
Le groupe des Agnatha est maintenant considéré comme polyphylétique. Il n'est pas montré sur cet arbre phylogénétique.
- Ici, les myxines sont considérées comme un clade séparé, comme dans l'article de Sweet & Donoghue de 2001 avec un arbre phylogénétique produit sans analyse cladistique[17]. Cependant, certains auteurs reconnaissent[29] que les myxines et les lamproies peuvent être plus proches les unes des autres dans leur propre clade, les Cyclostomata.
- Le clade des Proconodontida est aussi connu sous le nom de Cavidonti.
- Les Euconodonta sont aussi appelés "Conodonti" par Sweet & Donoghue[17], bien que ce nom ne soit pas beaucoup en usage.
Selon Terril et al., 2018[18], les conodontes sont à la base du groupe des cyclostomes:
| Vertebrata |
| |||||||||||||||||||||
Selon Bengston, 1983
▲
└─o Protoconodonta (éteint)
└─o Paraconodontida (éteint)
└─o Euconodonta (éteint)
Note: Les protoconodontes, fossiles du Cambrien, sont des éléments dont la composition est de nature organique ou faiblement phosphatique. Certains membres sont interprétés comme des crochets fossilisés de chaetognathes par Hubert Szaniawski sur la base d'une analyse comparative assez détaillée[30],[23].
▲
└─o Conodonta (éteint)
├─? Paraconodontida (éteint)
└─o Euconodonta (éteint)
├─o Proconodontida (éteint)
└─o Conodonti (éteint)
├─o Protopanderodontida (éteint)
└─o Prioniodontida (éteint)
Utilisation en stratigraphie
La systématique des conodontes est un bel outil bio-stratigraphique vu leur diversité et leur rapide évolution[31]. À ce niveau, ils priment facilement sur d'autres fossiles biostratigraphiques. On trouve de 0 à 1 000 conodontes (en moyenne) dans une roche d'un kilogramme (si celle-ci est susceptible d'accueillir ce fossile). Ils sont principalement étudiés en micro-paléontologie.
Pander Society
La Pander Society, fondée en 1967, est une société savante dédiée à l'étude des conodontes.
Elle doit son nom à Christian Heinrich von Pander (1794-1865), un embryologiste zoologue et paléontologue germano-balte, qui a employé le premier le terme de conodontes.
Notes et références
- (de) Christian Heinrich von Pander, Monographie des fossilen Fische des silurischen Systems des russisch-bamtischen Gouvernements, Buchdruckerei der Kaiserlichen Akademie der Wissenschaften, Saint-Pétersbourg, 1856, 91 pp. (Lire en ligne, Google books, Pages 1-112)..
- (en) Hass W.H., 1941. Morphology of Conodonts. Journal of Paleontology, Vol. 15, No. 1 (Jan., 1941), pp. 71-81 (lire en ligne sur JSTOR).
- (de) Eicheberg W., 1930. Conodonten aus dem Culm des Harzes. Paläontologische Zeitschrift, 1930, page 177-182, DOI:10.1007/BF03044446.
- (de) Schmidt H., 1934. Conodonten-Funde in ursprünglichem Zusammenhang. Paläontologische Zeitschrift, volume 16, numéros 1–2, pages 76–85, DOI:10.1007/BF03041668.
- (en) Scott H.W., 1934. The Zoological Relationships of the Conodonts. Journal of Paleontology, volume 8, numéro 4 (Dec., 1934), pages 448-455 (lire en ligne sur JSTOR)..
- (en) Scott H.W., 1942. Conodont Assemblages from the Heath Formation, Montana. Journal of Paleontology, volume 16, numéro 3, pages 293–300 (lire en ligne sur JSTOR).
- (en) Dunn D.L., 1965. Late Mississippian conodonts from the Bird Spring Formation in Nevada. Journal of Paleontology, volume 39, page 6 (lire en ligne).
- (en) Barnes C.R., 1967. A Questionable Natural Conodont Assemblage from Middle Ordovician Limestone, Ottawa, Canada. Journal of Paleontology; volume 41, numéro 6, pages 1557–1560 (lire en ligne sur JSTOR) .
- (en) Branson E.B. & Mehl M.G., 1931. Conodonts from Glassy Creek Shale of Missouri. Univ. Missouri Studies.
- (en) Hass W.H. & Lindberg M.L., 1946. Orientation of the Crystal Units of Conodonts. Journal of Paleontology, Vol. 20, No. 5 (Sep., 1946), pages 501-504.
- (en) Branson E.B. & Mehl M.G., 1941. New and Little Known Carboniferous Conodont Genera. Journal of Paleontology, Vol. 15, No. 2 (mars), pages 97-106.
- (en) Branson E.B. & Mehl M.G., 1944. Conodonts. in HW Shimer and RR Shrock, Index Fossils of North America.
- (en) Branson E.B., Mehl M.G. & Branson C.C., 1951. Richmond Conodonts of Kentucky and Indiana. Journal of Paleontology, Vol. 25, No. 1 (Jan.), pages 1-17.
- (en) Briggs D.E.G., Clarkson E.N.K. & Aldridge R.J., 1983. The conodont animal. Lethaia, Volume 16, Issue 1 (January 1983), Pages 1-14, DOI:10.1111/j.1502-3931.1983.tb01993.x.
- (en) Aldridge R.J., Briggs D.E.G., Smith M.P., Clarkson E.N.K. & Clark D.L., 1993. The anatomy of conodonts. Trans. R. Soc. Lond., 340, 405-421.
- (en) Fåhraeus L.E., 1983. Phylum Conodonta Pander, 1856 and Nomenclatural Priority. Systematic Zoology.
- (en) Sweet W.C & Donoghue P.C.J., 2001. Conodonts: Past, Present, Future. Journal of Paleontology, Vol. 75, No. 6, 75th Anniversary Issue (Nov., 2001), pages 1174-1184, DOI:10.1666/0022-3360(2001)075<1174:CPPF>2.0.CO;2 (lire en ligne sur JSTOR).
- (en) Terrill D.F., Henderson C.M. & Anderson J.S., 2018. New applications of spectroscopy techniques reveal phylogenetically significant soft tissue residue in Paleozoic conodonts†. J. Anal. At. Spectrom., 33, pages 992-1002, DOI:10.1039/C7JA00386B.
- (en) Donoghue P.C.J. & Rücklin M., 2014. The ins and outs of the evolutionary origin of teeth. Evolution & Development, 18(1), September 2014, DOI:10.1111/ede.12099.
- (en) Bergström S.M., Carnes J.B., Ethington R.L., Votaw R.B. & Wigley P.B., 1974. Appalachignathus, a New Multielement Conodont Genus from the Middle Ordovician of North America. Journal of Paleontology, volume 48, numéro 2, pages 227–235, JSTOR:1303249.
- (en) Purnell M.A. & Donoghue P.C.J., 1997. Architecture and functional morphology of the skeletal apparatus of ozarkodinid conodonts. Philosophical Transactions of the Royal Society B: Biological Sciences, volume 352, numéro 1361, pages 1545–1564, DOI:10.1098/rstb.1997.0141, PMC 1692076, Bibcode : 1997RSPTB.352.1545P.
- (en) Ferretti A., Malferrari D., Medici L. et al., 2017. Diagenesis does not invent anything new: Precise replication of conodont structures by secondary apatite. Sci Rep 7, 1624, DOI:10.1038/s41598-017-01694-4.
- (en) Henderson C.M., 2021. Conodonts. Encyclopedia of Geology (Second Edition), pages 435-445, DOI:10.1016/B978-0-08-102908-4.00113-2 (lien vers le chapitre).
- (en) Habermann D., Götte T., Meijer J., Stephan A., Richter A.D.K. & Niklas J.R., 2000. High resolution rare-earth elements analyses of natural apatite and its application in geo-sciences: Combined micro-PIXE, quantitative CL spectroscopy and electron spin resonance analyses. Nuclear Instruments and Methods in Physics Research Section B: Beam Interactions with Materials and Atoms, Volumes 161–163, March 2000, Pages 846-851, DOI:10.1016/S0168-583X(99)00998-2.
- (en) Anita G. Epstein, Jack Burton Epstein & Leonard Dorreen Harris, 1977. Conodont color alteration - an index to organic metamorphism. USGS Professional Paper, volume 995, pages 1–27, DOI:10.3133/pp995 (lire en ligne).
- (en) Königshof P., 2003. Conodont deformation patterns and textural alteration in Paleozoic conodonts: examples from Germany and France. Senckenbergiana Lethaea, volume 83, numéros 1/2, pages 149–156, DOI:10.1007/BF03043310 (Image).
- (en) Bengtson S., 1983. The early history of the Conodonta. Fossils and Strata 15, pages 5–19 (lire en ligne).
- (en) Azmi R.J. & Pancholi V.P., 1983. Early Cambrian (Tommotian) conodonts and other shelly microfauna from the Upper Krol of Mussoorie Syncline. Lesser Himalaya, with remarks on the Precambrian. Himalayan Geology, volume 11, pages 360-372.
- (en) Sarah J. Bourlat, Thorhildur Juliusdottir, Christopher J. Lowe, Robert Freeman, Jochanan Aronowicz, Mark Kirschner, Eric S. Lander, Michael Thorndyke, Hiroaki Nakano, Andrea B. Kohn, Andreas Heyland, Leonid L. Moroz, Richard R. Copley & Maximilian J. Telford, 2006. Deuterostome phylogeny reveals monophyletic chordates and the new phylum Xenoturbellida. Nature, volume 444, numéro 7115, pages 85–88, (ISSN 0028-0836), DOI:10.1038/nature05241, .
- (en) Szaniawski H., 2002. New evidence for the protoconodont origin of Chaetognaths. Acta Palaeontologica Polonica 47, pages 405–419 (lire en ligne).
- (en) Bassler R.S., 1925. Classification and stratigraphic use of the conodonts. Geological Society of America Bulletin, volume 36, issue 11, pages 218-220.
{kind=link}
Bibliographie
- (en) Supplement 2 : Conodonta (Clark D.L., Sweet W.C., Bergström S.M., sous la direction de Richard A. Robison, 1981, (ISBN 0813730287)). dans Treatise on invertebrate paleontology, Part W: Miscellanea : Conodonts Conoidal Shells of Uncertain Affinities, Worms, Trace Fossils, and Problema - Geological Society of America (sous la direction de Raymond C. Moore).
- (en) Conodonts. Maurits Lindström, Elsevier Publishing Company, 1964, 196 pages (ISBN 0444403728).
- (en) Simon J. Knell, 2014. The great fossil enigma: the search for the conodont animal. Earth Sciences History, volume 33, numéro 1, pages 179-181 (lire en ligne sur JSTOR).
Liens externes
- Ressources relatives au vivant :
- (cs + en) BioLib
- (en) Fossilworks
- (en) Interim Register of Marine and Nonmarine Genera
- (en) ZooBank
 Portail de la paléontologie
Portail de la paléontologie  Portail de l’ichtyologie
Portail de l’ichtyologie