Coiffe (biologie)
La coiffe ou 5'-cap est un nucléotide modifié que l'on trouve à l'extrémité 5' des ARN messagers dans les cellules eucaryotes. C'est une modification co-transcriptionnelle qui est introduite par l'action successive de plusieurs enzymes localisées dans le noyau. La coiffe joue plusieurs rôles, elle protège en particulier les ARN d'une dégradation enzymatique par des ribonucléases et, après son export dans le cytoplasme, elle permet le recrutement du ribosome qui va traduire l'ARN messager.
La coiffe est un élément essentiel permettant aux ARN messagers d'être traduits dans la cellule. Ce processus nécessite l'action de plusieurs protéines et en particulier des facteurs d'initiation de la traduction. L'un d'entre eux, eIF4E, se fixe spécifiquement sur la coiffe.
Structure
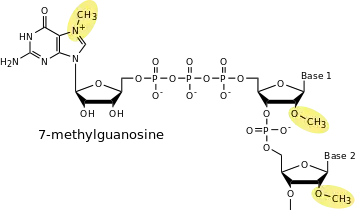
La coiffe se compose d'une guanosine méthylée en position N7, reliée au premier nucléotide transcrit par une liaison 5'-5' triphosphate. Les riboses des deux premiers nucléotides de l'ARN transcrit sont également méthylés sur leur position 2'-hydroxyle[1]. La liaison 5'-5' est très inhabituelle, et fait que, après modification, l'extrémité 5' de l'ARN ressemble chimiquement à son extrémité 3'. Les deux nucléotides terminaux portent en effet un cis-diol (2'-OH et 3'-OH). On note parfois la structure de la coiffe 7mGpppN.
La méthylation sur le 2'-OH du ribose des deux premiers nucléotides peut être variable en fonction des types cellulaires et des espèces. On trouve des coiffes non méthylées, mono-méthylées sur la position 1 et di-méthylées sur les positions 1 et 2[2]. Lorsque le premier nucléotide transcrit est un A, celui-ci peut subir une méthylation additionnelle sur la position N6 de l'adénine.
Synthèse
La synthèse de la coiffe est un processus en plusieurs étapes qui se déroule dans le noyau et implique différentes enzymes. L'ARN transcrit par l'ARN polymérase II commence par un nucléotide 5' triphosphate. L'extrémité triphosphate est d'abord clivée par une triphosphatase (EC 3.1.3.33) qui libère un 5' diphosphate :
pppN1pN2p... → ppN1pN2p... + Pi
Ensuite une guanylyltransférase (EC 2.7.7.50) forme la liaison 5'-5' triphosphate, en utilisant le GTP comme donneur. Ceci libère une molécule de pyrophosphate provenant de l'hydrolyse du GTP.
Gppp + ppN1pN2p... → GpppN1pN2p... + PPi
Enfin, des méthyltransférases ajoutent les différents groupements méthyle sur le N7 du G (EC 2.1.1.56) et les nucléotides N1 et N2. Le donneur de méthyle est la S-adénosylméthionine.
Suivant les espèces, certaines de ces différentes activités enzymatiques peuvent être regroupées sur une ou plusieurs protéines multifonctionnelles[3]. Chez la levure, les activités triphosphatase, guanylyltransférase et N7-méthyltransférases sont portées par des protéines distinctes. Chez les mammifères et en particulier chez l'homme, la triphosphatase et la guanylyltransférase sont portées par la même protéine, Hcmlp. La synthèse de la coiffe est un processus co-transcriptionnel, qui se produit aussitôt que la chaîne d'ARN naissante émerge de l'ARN polymérase II. Les enzymes de synthèse de la coiffe sont en particulier associées au domaine C-terminal de la polymérase[4].
Les différents gènes codant les enzymes impliqués dans la synthèse de la coiffe des ARN sont des gènes essentiels, dont l'inactivation est létale pour la cellule[5].
Fonction
La coiffe des ARN synthétisés par l'ARN polymérase II, et en particulier les ARN messagers, joue plusieurs rôles cellulaires essentiels :
- Stabilisation : La coiffe bloque en particulier l'action des exonucléases 5' → 3' et augmente la durée de vie des ARN messagers; elle neutralise la détection des ARNm viraux par les mécanismes de l’immunité innée[6]
- Export nucléaire : La coiffe est reconnue dans le noyau par un complexe de fixation de la coiffe (CBC ou cap-binding complex). Ce complexe intervient en particulier dans l'export de l'ARNm maturé au niveau du pore nucléaire;
- Traduction en protéine : Dans le cytoplasme, la coiffe se lie au facteur d'initiation eIF4E. eIF4E forme un complexe avec eIF4G qui recrute le ribosome via eIF3. La traduction d'un ARNm cellulaire est donc, dans la grande majorité des cas, dépendante de la présence d'une coiffe en 5'.
Catabolisme
Les ARN messagers subissent un "décoiffage" qui permet leur recyclage. Il existe des enzymes spécifiques permettant de réaliser cette étape, chez la levure, c'est la protéine Dcp1p qui catalyse cette étape sur les ARN entiers en coupant la liaison 5'-5' triphosphate[7]. Le produit du décoiffage est un ARN 5'-monophosphate et du 7-méthyle GDP. Il existe une autre enzyme de recyclage, DcpS, capable de couper la coiffe seule (le dinucléotide 7mGpppN, clivé de l'ARN messager), qui libère du 7-méthyle GMP et un nucléotide diphosphate.
Notes et références
- (en) Reddy R., Ro-Choi T.S., Henning D., Busch H., « Primary sequence of U-1 nuclear ribonucleic acid of Novikoff hepatoma ascites cells. », J. Biol. Chem., vol. 249, no 20, , p. 6486-6494 (PMID 4370679)
- (en) Banerjee A.K., « 5'-terminal cap structure in eucaryotic messenger ribonucleic acids. », Microbiol. Rev, vol. 44, no 2, , p. 175-205 (PMID 6247631)
- (en) Shuman S., « Structure, mechanism, and evolution of the mRNA capping apparatus. », Prog. Nucleic Acid Res. Mol. Biol., vol. 66, , p. 1-40 (PMID 11051760)
- (en) McCracken S., Fong N., Rosonina E., Yankulov K., Brothers G., Siderovski D., Hessel A., Foster S., Shuman S., Bentley D.L., « 5'-Capping enzymes are targeted to pre-mRNA by binding to the phosphorylated carboxy-terminal domain of RNA polymerase II. », Genes Dev., vol. 11, , p. 3306-3318 (PMID 9407024)
- (en) Schwer B., Shuman S., « Mutational analysis of yeast mRNA capping enzyme. », Proc. Natl. Acad. Sci. USA, vol. 90, , p. 4228-4232 (PMID 8183907)
- medecinesciences.org
- (en) Beelman C.A., Stevens A., Caponigro G., LaGrandeur T.E., Hatfield L., Fortner D.M., Parker R., « An essential component of the decapping enzyme required for normal rates of mRNA turnover. », Nature, vol. 382, no 6592, , p. 642-646 (PMID 8757137)
Articles connexes
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire