Cellule de Schwann
Les cellules de Schwann (ou neurolemmocytes) sont une variété de cellules gliales qui assurent principalement l'isolation myélinique des axones du système nerveux périphérique des chordés (on les classe donc parmi les « cellules gliales périphériques »). Comme les oligodendrocytes du système nerveux central, elles assurent la myélinisation—c'est-à-dire l'isolation électrique—des axones mais dans le système nerveux périphérique. Il existe néanmoins de petites différences entre ces deux types de cellules.
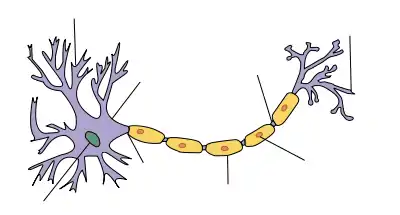

Leur nom est un hommage au physiologiste allemand Theodor Schwann.
Histologie
Les cellules de Schwann n'existent qu'au niveau du système nerveux périphérique, alors que les oligodendrocytes jouent ce rôle dans le système nerveux central ; elles constituent la gaine de myéline autour d'un seul axone, alors que les oligodendrocytes peuvent myéliniser plusieurs axones.
Un réseau bien développé de cellules de Schwann est en forme de barres de feuille de papier, avec des couches de myéline entre deux bobines. Les couches intérieures de l'emballage, qui sont essentiellement des matériaux de membrane, forment la gaine de myéline alors que la couche supérieure du cytoplasme nucléé forme les neurolemmes. Seul un petit volume de résidus cytoplasmique communique de l'intérieur vers les couches extérieures, il s'agit de l'incisure de Schmidt Lanterman. On peut également observer une communication cellulaire, par l'intermédiaire de jonctions à interstices (jonctions gap) : l'ensemble des cellules de Schwann forment ainsi un Syncytium fonctionnel.
Une seule cellule de Schwann recouvre au mieux un millimètre de longueur d'axone. Il en faut donc des centaines ou milliers pour couvrir entièrement un axone, (qui peut parfois atteindre la longueur d'un corps).
L'écart existant entre deux cellules de Schwann (secteur non couverts par la gaine de myélin) est dit Nœud de Ranvier. Ces nœuds constituent d'importants sites d'échanges ioniques entre l'axone et les liquides extracellulaires (on en trouve notamment dans le système nerveux somatique).
Cet agencement permet la conduction saltatoire du potentiel d'action à grande vitesse (le PA « saute » par-dessus la myéline ce qui donne une vitesse parfois supérieure à cent mètres par seconde), ainsi que des économies d'énergie.
Développement des cellules de Schwann
Les cellules de Schwann dérivent d'une structure dite crête neurale, qui se forme au-dessus du tube neural lors du développement embryonnaire. Lorsqu'elles sont en contact avec des axones, les cellules de la crête neurale se différencient en « précurseur de cellule de Schwann ». Elles deviennent ensuite des « cellules de Schwann immatures », qui entourent alors des dizaines d'axones à la fois. Elles peuvent ensuite devenir des cellules de Schwann myélinisantes et ne protéger qu'un seul axone, ou alors devenir des cellules de Schwann non-myélinisantes et entourer plusieurs axones de faible diamètre. Ces variations semblent guidées par le diamètre des axones avec lesquels elles sont en contact. Les axones les plus larges induisent la différenciation en cellule de Schwann promyélinisante par le biais d'un signal moléculaire ; alors que les axones plus fins n'émettent pas assez de signal pour induire cette différenciation. De même, l'épaisseur de la gaine de myéline sera proportionnelle au diamètre de l'axone. Dans une fibre myélinisée, le manchon de myéline agit comme un isolant. Les inversions de charges sautent d'un nœud de Ranvier (espacement entre chaque gaine de myéline) à l'autre, permettant à l'influx nerveux de se propager plus vite. La vitesse de propagation du potentiel d'action le long du neurone est en moyenne de 50 m/s, mais cette vitesse varie selon différents facteurs tels que la température, le diamètre et s'il y a de la myéline ou non.
Rôle de la myélinisation
Le système nerveux des vertébrés s'appuie sur cette gaine de myéline, pour l'isolation et comme une méthode de diminution de la capacité des membranes dans l'axone, ce qui permet à la conduction saltatoire de se produire et une augmentation de la vitesse d'impulsion, sans augmentation de diamètre axonal.
La gaine de myéline étant isolante, les canaux sodiques voltage-dépendants (responsables de la conduction du potentiel d'action) ne se trouvent, sur un axone myélinisé, qu'au niveau des zones non recouvertes de myéline (cône axonal et nœuds de Ranvier donc).
Les « cellules non myélinisantes de Schwann » sont impliquées dans l'entretien des axones neuronaux ; elles sont cruciales pour la survie.
Implication en pathologie et en thérapie
Les cellules de Schwann sont impliquées dans diverses pathologies du système nerveux périphérique, dont :
- Maladie de Charcot-Marie-Tooth, une neuropathie héréditaire sensitivo-motrice. La myélinisation est diminuée dans certains cas de cette pathologie, ce qui a pour effet de diminuer la vitesse de conduction de l'influx nerveux le long des nerfs périphériques.
- Sclérose en plaques : des études expérimentales ont depuis 2001 implanté les cellules de Schwann pour induire la remyélinisation chez les patients touchés cette maladie, car connues pour leur rôle de soutien à régénération nerveuse. Néanmoins, les cellules de Schwann interagissent trop fortement avec les astrocytes du système nerveux central, ce qui les empêche d'atteindre les lésions et de remyéliniser efficacement les neurones touchés. Les recherches s'orientent donc de plus en plus vers des précurseurs de cellules de Schwann ou des cellules apparentées comme les Olfactory Ensheating Cells (cellules d'engainage des nerfs olfactifs (en) qui réagissent beaucoup moins avec les populations gliales du système nerveux central).
- Cancer : lors d'une expérience récente (publication 2016) sur les effets de l'exposition aux ondes de téléphones portables, une tumeur rare des cellules de Schwann du cœur a touché 2 à 6 % des rats mâles exposés mais pas les femelles ni aucun des rongeurs du groupe témoin (non-exposés au rayonnement)[1].
Notes et références
- Michael Wyde & al. (2016) « Report of Partial findings from the National Toxicology Program Carcinogenesis Studies of Cell Phone Radiofrequency Radiation in Hsd: Sprague Dawley® SD rats (Whole Body Exposure) » doi: https://dx.doi.org/10.1101/055699 ; rapport provisoire, déposé sur le serveur de prépublication « bioRxiv preprint » le 26 mai 2016 dans le cadre du National Toxicology Program (NTP), publié en licence cc-by-4.0 (résumé et version PDF, 74pp
Voir aussi
Articles connexes
Bibliographie
 Portail des neurosciences
Portail des neurosciences  Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire