Caenorhabditis elegans
Caenorhabditis elegans est un petit ver d'un millimètre environ, transparent et non-parasitaire. C'est un nématode de la famille des Rhabditidae.
| Règne | Animalia |
|---|---|
| Embranchement | Nematoda |
| Classe | Secernentea |
| Ordre | Rhabditida |
| Famille | Rhabditidae |
| Genre | Caenorhabditis |
Sous-espèces de rang inférieur
Le nom est formé des deux racine grecques caeno- (καινός (caenos), récent), et rhabditis (ῥάβδος (rhabdos), tige, bâton) et du mot latin elegans (élégant). En 1900, Maupas nomma l'espèce Rhabditides elegans, puis Osche la plaça dans le sous-genre Caenorhabditis en 1952, et en 1955, Dougherty établit Caenorhabditis au rang de genre.[5]
Ce ver est un organisme modèle en biologie, car il permet l'étude, entre autres, de l'apoptose (mort programmée de certaines cellules), du développement embryonnaire et du vieillissement.
Description

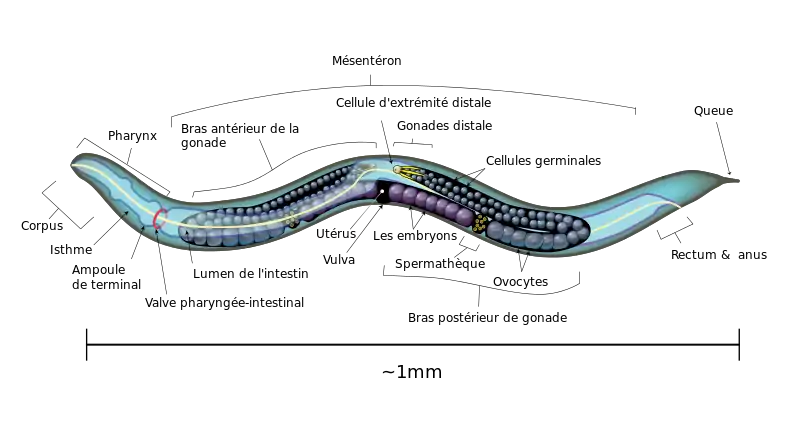
C. elegans est un petit ver transparent d'environ un millimètre de longueur, se nourrissant principalement de bactéries. Son embryogenèse ne dure que 16 heures et peut être facilement observée in vitro[6]. La plupart des individus sont hermaphrodites (XX) et produisent à la fois des œufs et des spermatozoïdes. Des mâles (XO) peuvent aussi être produits par non-disjonction du chromosome à la méiose ou lors de croisements entre mâles et hermaphrodites. Les spermatozoïdes des mâles ne possèdent pas de flagelles et utilisent un mode de déplacement amiboïde[7]. Ce nématode est capable de se reproduire après trois jours de développement depuis l'œuf, et sa durée de vie est d'environ trois semaines[8].
Pour l'observation au microscope photonique, il peut être observé vivant entre lame et lamelle sur de la gélose ou fixé entre lame et lamelle.
Anatomie
Vision des couleurs
C. elegans ne dispose d'aucun organe visuel, ni même de gènes codant une opsine. Il est néanmoins capable de distinguer certaines couleurs de son environnement, ce qui lui permet d'éviter les zones riches en bactéries toxiques. Cette vision des couleurs est due à des gènes codant des protéines sensibles aux contraintes, jkk-1 (une kinase) et lec-3 (une lectine)[9],[10].
Animal-modèle de laboratoire

C. elegans a été introduit dans les laboratoires de génétique à la fin des années 1960 par Sydney Brenner, pour répondre au besoin d'un organisme modèle génétique pour comprendre l'élaboration d'un organisme pluricellulaire et la neurobiologie. Les « pères » du modèle Caenorhabditis elegans, Sydney Brenner, John Sulston et Robert Horvitz, ont reçu le prix Nobel en 2002[11].
Le nématode possède un nombre quasiment fixe de cellules (eutélie). En 1983, Sir John Sulston en publia les lignages cellulaires, c'est-à-dire toute la généalogie des cellules depuis l'œuf jusqu'à l'adulte, quasi-invariant d'un individu à l'autre.
- L'adulte hermaphrodite (99,5 % des individus) est composé de 959 cellules somatiques dont le lignage a pu être défini (certaines de ces cellules fusionnent entre elles à un moment du développement, formant des syncytiums).
- L'adulte mâle est composé de 1031 cellules.
Au cours de la vie du ver, il y a multiplication cellulaire, et mort par apoptose de 131 cellules (1 090 cellules sont produites et il en reste 959 chez l'adulte). Chacune de ces 131 cellules se « suicide » à peu près au même moment et au même endroit dans tous les embryons.
La simplicité de ce ver s'est imposée dans l'étude du génome, de la régulation de l'expression des gènes, de la polarité cellulaire, de l'apoptose, de l'organogenèse et du vieillissement. Une partie des cellules du ver (302) sont des cellules neuronales, permettant également l'étude du système nerveux et du comportement.
Génétique et épigénétique
Génome
En 1998, ce ver fut le premier animal dont le génome ait été totalement séquencé : 97 millions de paires de bases réparties en six paires de chromosomes (organisme diploïde) codant 19 099 gènes. Quarante pour cent d'entre eux auraient des équivalents dans le patrimoine génétique humain. Les ancêtres de ce ver ainsi que ceux des hommes auraient divergé il y a 550 millions d'années. L'ensemble des informations génomiques concernant cet organisme sont compilées dans WormBase.
Épigénétique
En 2017 Klosin et al. ont montré qu'un stress thermique peut induire chez C. elegans l'expression de copies réprimées (de manière endogène) de gènes - parfois appelés ADN non codant ou "junk DNA", avec un effet persistant pour au moins 14 générations de vers. La chromatine est modifiée en début d'embryogénèse, avant le début de la transcription et la modification est héritée par les ovules comme par les spermatozoïdes. Dans ce cas, on observe une triméthylation altérée de l'histone H3 lysine 9 (H3K9me3) avant le début de la transcription zygotique[13].
Mort programmée
Robert Horvitz et son équipe de Boston ont pu identifier par mutagenèse au hasard les divers gènes produisant les protéines qui contrôlent la mort cellulaire programmée durant le développement de C. elegans. Ils ont trouvé des mutants chez lesquels les 131 cellules normalement destinées à disparaître par apoptose échappent à la mort programmée.
Les gènes intervenant dans les mutations ont été appelés ced (pour cell death-abnormal). Les 131 cellules qui meurent généralement survivent chez les vers dont le gène ced-3 ou ced-4 a été inactivé. Le gène ced-9 a l'effet inverse puisque l'absence de la protéine CED-9 entraîne une augmentation de l'apoptose des cellules de l'embryon.
L'étude génétique et moléculaire de ces mutations[14] a montré que les protéines CED-3 et CED-4 sont responsables du déclenchement de l'apoptose et que ced-9 restreint leur effet. Si les trois gènes ced-3, ced-4 et ced-9 sont inactivés, aucune cellule de l'embryon ne subit de mort programmée. La survie des 959 cellules de l'embryon tient donc à un jeu d'équilibre entre les exécuteurs (ced-3, ced-4) et le protecteur (ced-9). Suivant la formulation de Nicole Le Douarin[14] « chaque cellule de l'embryon produit les protéines capables de la tuer. Ne survivent que celles qui sont aptes, pour un temps, à s'opposer à la mise en œuvre du programme de mort ».
Un troisième gène provoquant la mort programmée a été découvert en 1998 par l'équipe de Robert Horvitz. Il s'agit de egl-1 (pour egg-laying defective) qui avec ced-3 et ced-4 est nécessaire pour réaliser le développement normal de C. elegans. La mutation d'un de ces gènes se traduit par la survie de presque toutes les cellules normalement vouées à disparaître. La protéine egl-1 se lie à celle de ced-9 pour lui interdire son rôle de protecteur.
Les études génétiques de C. elegans ont conduit à l'identification de plus de deux douzaines de gènes dans la spécification des cellules destinées à mourir ou à vivre, dans l'activation du programme de suicide puis l'élimination des cellules mortes (Conradt & Xue[15], 2005). Mais ced-3, ced-4, ced-9 et egl-1 paraissent bien être les acteurs les plus importants du programme de mise à mort.
La mort de certaines cellules embryonnaires se présente dès lors comme un processus naturel activé en certains lieux, à certains moments, par l'expression du gène egl-1.
Des équivalents de ces gènes chez l'homme ont été trouvés : bax,bad,bak et bim pour l'apoptose, et bcl-2,bclxl pour la survie de cellules. Les protéines effectrices de la voie apoptotique sont des enzymes appelées caspases. Ainsi, la protéine produite par ced-3 est une caspase.
Résistance
Comme de nombreux nématodes, à certains stades de son développement, ce ver peut produire des larves dauer, plus résistantes et capables de survivre durant plusieurs mois en diapause.
On a retrouvé des spécimens d'étude de C. elegans vivants dans les débris de la navette spatiale Columbia après son accident en février 2003[16].
Notes et références
- Les modalités de la reproduction et le déterminisme du sexe chez quelques nematodes libres. Nigon V. Ann. Sci. Nat. Zool. Biol. Anim. 1949;11:1–132.
- Moerman DG, Waterston RH Spontaneous unstable unc-22 IV mutations in C. elegans var. Bergerac. Genetics, volume 108, issue 4, pages 859–77, Décembre 1984, , pmc 1224270
- Babity J. M. Tc1 transposition and mutator activity in a Bristol strain of Caenorhabditis elegans|journal=Molecular & General Genetics : Mgg volume 222, issue 1, pages 65–70
- Harris L. J. Structural analysis of Tc1 elements in Caenorhabditis elegans var. Bristol (strain N2). Plasmid, volume= 22; issue 1, pages=10–21, 1989,
- H Ferris, « Caenorhabditis elegans » [archive du ], University of California, Davis, (consulté le )
- Scott-F Gilbert, Susan-R Singer, Sylvie Rolin (trad), Etienne Brachet (trad.), Biologie du développement, De Boeck,
- Xuan Ma, Yanmei Zhao, Wei Sun et Katsuya Shimabukuro, « Transformation: how do nematode sperm become activated and crawl? », Protein & Cell, vol. 3, no 10, , p. 755–761 (ISSN 1674-800X, PMID 22903434, PMCID 4875351, DOI 10.1007/s13238-012-2936-2, lire en ligne, consulté le )
- (en) L. Byerly, R.C. Cassada et R.L. Russell, « The life cycle of the nematode Caenorhabditis elelgans : I. Wild-type growth and reproduction », Developmental Biology, vol. 51, no 1, , p. 23-33
- (en) Lauren A. Neal1 et Leslie B. Vosshall, « Eyeless worms detect color », Science, vol. 371, no 6533, , p. 995 (DOI 10.1126/science.abg5201).
- (en) D. Dipon Ghosh, Dongyeop Lee, Xin Jin, H. Robert Horvitz et Michael N. Nitabach, « C. elegans discriminates colors to guide foraging », Science, vol. 371, no 6533, , p. 1059-1063 (DOI 10.1126/science.abd3010).
- Nobelprize.org, « The Nobel Prize in Physiology or Medicine 2002 », (consulté le )
- Greer E, Maures T, Ucar D, et coll. Transgenerational epigenetic inheritance of longevity in Caenorhabditis elegans. Nature 2011, 479:365-71.
- Adam Klosin1, Eduard Casas, Cristina Hidalgo-Carcedo, Tanya Vavouri & Ben Lehner (2017) Transgenerational transmission of environmental information in C. elegans, Science News, 21 avril 2017:Vol. 356, Issue 6335, pp. 320-323 DOI: 10.1126/science.aah6412
- Nicole Le Douarin, Les cellules souches, porteuses d'immortalité, Odile Jacob,
- Barbara Conradt, Ding Xue, « Programmed cell death », WormBook, ed. The C. elegans Research Community, WormBook, (DOI 10.1895/wormbook.1.32.1)
- (en) « Worms survived Columbia disaster », BBC News, (lire en ligne)
Voir aussi
Articles connexes
 Portail de la zoologie
Portail de la zoologie  Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire