Apis mellifera
Abeille européenne, Avette, Mouche à miel

| Règne | Animalia |
|---|---|
| Embranchement | Arthropoda |
| Classe | Insecta |
| Ordre | Hymenoptera |
| Sous-ordre | Apocrita |
| Super-famille | Apoidea |
| Famille | Apidae |
| Sous-famille | Apinae |
| Tribu | Apini |
| Genre | Apis |
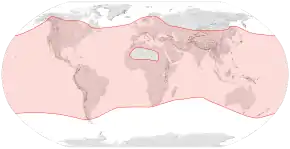
Répartition géographique
Abeille européenne, avette ou mouche à miel (Apis mellifera) est une abeille à miel originaire d'Europe. Elle est considérée comme semi-domestique. C'est une des abeilles élevées à grande échelle en apiculture pour produire du miel et pour la pollinisation.
Le nom de genre Apis est le terme latin signifiant « abeille », et l'épithète spécifique mellifera signifie « qui donne du miel », en référence à la tendance de l'espèce à produire une grande quantité de miel pour le stocker au printemps et en été avant consommation pendant l'hiver.
L'abeille européenne occupe désormais tous les continents sauf l'Antarctique. En raison de sa grande diffusion, cette espèce est le pollinisateur le plus important pour l'agriculture à l'échelle mondiale. Un certain nombre de ravageurs et de maladies menacent l'abeille, en particulier le syndrome d'effondrement des colonies[1].
Sous-espèces
.jpg.webp)
Apis mellifera comporte vingt-huit sous-espèces (toutes fécondables entre elles) réparties en cinq groupes majeurs (selon leurs évolutions et leurs répartitions géographiques) basés sur le travail de Friedrich Ruttner et confirmés par l'analyse de l'ADN mitochondrial :
- Lignée ou type A (groupe africain)
- Apis mellifera adansonii (Latreille)
- Apis mellifera capensis (Eschscholtz)
- Apis mellifera intermissa (Maa)
- Apis mellifera litorea (Smith)
- Apis mellifera monticola (Smith)
- Apis mellifera sahariensis (Baldensperger)
- Apis mellifera scutellata (Lepeletier)
- Apis mellifera sicula (Montagano)
- Apis mellifera unicolor (Latreille)
- Lignée ou type C (groupe Carniole, de l'Europe de l'Est et du sud des Alpes)
- Apis mellifera carnica (Pollman)
- Apis mellifera cecropia (Kiesenwetter)
- Apis mellifera ligustica (Spinola)
- Apis mellifera macedonica (Ruttner)
- Lignée ou type M (groupe méditerranéen, de l'Europe de l'Ouest et du Nord)
- Apis mellifera iberica (Engel)
- Apis mellifera mellifera (Linnaeus)
- Apis mellifera siciliana (Grassi)
- Lignée ou type O (groupe du Moyen-Orient et de l'Asie centrale)
- Apis mellifera adami (Ruttner)
- Apis mellifera anatoliaca (Maa)
- Apis mellifera armeniaca (es)
- Apis mellifera caucasia (Pollmann)
- Apis mellifera cypria (Pollman)
- Apis mellifera lamarckii (Cockerell)
- Apis mellifera meda (Skorikov)
- Apis mellifera syriaca (Skorikov)
- Apis mellifera taurica (Alpatov)
- Lignée ou type Y[2] (groupe Nord-Est, de l'Éthiopie et du Yemen )
- Apis mellifera jemenitica (Ruttner)
- Reste à classer
- Apis mellifera artemisia (Engel)
- Apis mellifera remipes (Gerstaecker)
- Apis mellifera ruttneri (Sheppard, Arias, Grech & Meixner)
- Apis mellifera sossimai (Engel)
Les plus utilisées en apiculture
Les variétés les plus utilisées en apiculture sont[3] :
- La Buckfast, un hybride de multiples variétés sélectionnées
- Apis mellifera ligustica, l'abeille jaune Italienne
- Apis mellifera carnica, l'abeille carniolienne
- Apis mellifera mellifera, l'abeille noire
- Apis mellifera caucasica, l'abeille du Caucase
Identification des sous-espèces
Une analyse biométrique détaillée d'une trentaine d'abeilles d'une même colonie est un moyen assez fiable d'identifier la variété dont on dispose. On doit alors étudier la coloration de l'abdomen, la pilosité abdominale, la largeur du tomentum (bande pileuse abdominale), la longueur de la langue (proboscis), l'indice cubital et la transgression discoïdale des nervures de l'aile. Cette analyse nécessite de disposer d'un microscope ou de passer par des sites Internet spécialisés[4] qui permettent de faire ces analyses à partir d'images d'ailes envoyées par les utilisateurs. Les résultats sont à comparer aux grilles de références établies par Ruttner[5].
Une analyse éthologique est moins précise mais permet de se faire une idée sans matériel particulier. On doit alors tenir compte de critères tels que :
- la fécondité,
- l'ardeur à butiner,
- la résistance du couvain et des abeilles aux maladies courantes,
- la lenteur à l'essaimer,
- la résistance aux intempéries,
- l'ardeur à construire,
- la douceur,
- la tenue sur cadre,
- la propension plus ou moins importante à propoliser,
- l'apparence des rayons (droit ou pas),
- le sens de l'orientation
Puis comparer les résultats obtenus au tableau de caractéristiques créé par Frère Adam (Karl Kehrle)[6]. Certaines variétés comme les Buckfast (assez pures) sont toutefois assez reconnaissables notamment pour leurs bourdons très jaune doré voire blonds.
Description
.jpg.webp)






La Reine mesure de 15 à 20 millimètres de long et les ouvrières entre 11 et 13 millimètres. Cependant certaines sous-espèces des climats plus chauds de l'Afrique sont plus petites. On peut facilement reconnaître la reine à sa taille et son abdomen élargi.
La couleur de base de l'abeille occidentale est brune, avec les premiers segments abdominaux jaune, orange à rouge. Le thorax poilu est jaune brunâtre. Contrairement à une croyance populaire, l'abdomen de l'abeille n'est pas coloré en noir et jaune. Les illustrations colorées dans les livres pour enfants, par exemple, correspondent en fait à la coloration abdominale jaune et noir de la guêpe.
Les pattes des abeilles sont structurées comme celles des autres insectes. Les pattes postérieures des ouvrières servent à recueillir le pollen. Le bas des pattes est recouvert de longs poils formant une sorte de « panier » à pollen.
L'abeille, outre les mandibules, dispose d'un tuyau d'aspiration.
Biologie et cycle de vie
En zone tempérée, les abeilles survivent à l'hiver en tant que colonie, et la reine, seule femelle féconde, commence à pondre des œufs à la fin de l'hiver (probablement déclenché par la longueur du jour). La reine pond les œufs dont toutes les autres abeilles sont produites. Sauf pour une brève période (quand elle peut voler pour s'accoupler avec des mâles ou partir plus tard avec un essaim pour établir une nouvelle colonie), la reine quitte rarement la ruche après que les larves sont devenues des abeilles.
Le processus de naissance est ensuite variable selon qu'il s'agit d'une reine, d'une ouvrière ou d'un faux bourdon.
| Reine | Ouvrière | Mâle | |
|---|---|---|---|
| Œuf | fécondé | fécondé | non fécondé |
| Stockage des œufs | Cellule royale | Alvéole classique | Alvéole de mâles (plus grosses et rebondies) |
| Alimentation | Gelée royale | Gelée royale les 3 premiers jours puis alimentation composée |
Gelée pour mâles |
| Temps de développement global | 16 jours | 21 jours | 24 jours |
| • Œuf | • 3 jours | • 3 jours | • 3 jours |
| • Larve | • 5,5 jours | • 6,5 jours | • 7 jours |
| • Nymphe | • 8 jours | • 12 jours | • 14 jours |
| Poids à l'éclosion | environ 200 mg | environ 100 mg | environ 200 mg |
| Longueur du corps | 18–22 mm | 12–15 mm | 15–17 mm |
| Maturité sexuelle | environ 7 jours | environ 14 jours | |
| Durée de vie | Jusqu'à 7 ans | en été 2 à 6 semaines
en hiver 4 à 7 mois |
1–3 mois |
- Jour J : la reine dépose chaque œuf (bâtonnet blanc d'environ 1,5 mm de long et 0,4 mm de diamètre, pesant 0,13 mg) dans une cellule préparée par des abeilles ouvrières.
- J + 3 : L'œuf donne une petite larve nourrie par des abeilles «nourrices» (abeilles ouvrières qui maintiennent l'intérieur de la colonie).
- J 3 à J 8 ou 10 : C'est le stade larvaire. Il dure 6 jours pour une reine et 7 jours pour une ouvrière.
- J 9 ou J 10 : Operculation de la cellule. Les larves sont scellées dans leur cellule par les abeilles infirmières et commencent leur stade nymphal (7 jours pour les reines, 12 pour les ouvrières, 14 pour les mâles)
- J + 16 : l'imago royal apparaît et peut devenir la nouvelle reine.
- J+20 à J+24 : Vol nuptial de la reine. Début de la ponte : 3 jours après fécondation.
- J + 21 : la larve d'ouvrières apparaît comme une abeille adulte et se met immédiatement au travail.
- J + 25 : la larve mâle donne naissance à un faux-bourdon.
Les éleveurs de reine considèrent qu'une bonne nutrition pendant la phase larvaire (et une bonne génétique) est cruciale pour la qualité de la reine. Pendant les stades larvaire et pupal, les parasites peuvent endommager (ou détruire) la pupe ou la larve.
La population moyenne d'une ruche saine en plein été varie de 40 000 à 80 000 abeilles. Elle est 10 fois inférieure en hiver.
Les cadres d'une ruche contenant les larves et les nymphes sont appelées « cadres de couvain », et sont vendus aux apiculteurs accompagnés d'au moins un cadre de pollen et un cadre de miel (avec des abeilles nourricières) pour commencer de nouvelles ruches.
Périodiquement, la colonie détermine qu'une nouvelle reine est nécessaire. Il y a trois causes générales:
- La ruche est remplie de miel, laissant peu de place pour de nouveaux œufs. Cela déclenchera un essaimage, où la vieille reine prendra environ la moitié des abeilles ouvrières pour fonder une nouvelle colonie et laisser la nouvelle reine avec l'autre moitié des ouvrières pour continuer l'ancienne.
- La reine vieillit et commence à manquer de spermatozoïdes et à avoir des problèmes de communication (diminution des phéromones de la reine dans toute la ruche). C'est ce que l'on appelle la supersédure (remplacement), et à la fin de la supersedure, la vieille reine est généralement tuée.
- La vieille reine meurt soudainement, une situation connue sous le nom de supersédure d'urgence. Les ouvrières trouvent plusieurs œufs (ou larves) de la tranche d'âge appropriée et tentent de les développer en reines. La supersédure d'urgence peut généralement être reconnue parce que les nouvelles cellules de la reine sont construites à partir de cellules du rayon, au lieu d'être suspendues sous le haut d'un cadre.
Castes sociales
Comme toutes les abeilles mellifères, l'abeille européenne est eusociale, créant des colonies avec une seule femelle féconde (ou « reine »), de nombreuses femelles stériles ou « ouvrières », et une faible proportion de mâles fertiles ou « faux-bourdon ». Chaque colonie peut héberger des dizaines de milliers d'abeilles. Les activités de colonie sont organisées par une communication complexe entre les individus, à travers les odeurs et une langue dansée.
La reine


La reine se distingue des autres abeilles par sa plus grande taille (1,5 à 2 cm contre 1,1 et 1,3 cm). Son abdomen et ses organes génitaux sont plus développés. Elle passe tout son temps à pondre, et c'est elle qui donne naissance à toutes les abeilles d'une ruche.
En pleine saison, la reine peut pondre plus de 2 500 œufs par jour (plus que sa masse corporelle) durant une miellée d'été et à l'apogée de sa fertilité (à l'âge de deux ans). Elle produit deux types d'œufs: les œufs fécondés, qui donnent naissance aux abeilles femelles (ouvrières ou reines), et les œufs non fécondés d'où sortent les abeilles mâles (faux bourdons).
Les reines ne sont pas élevées dans les cellules de ponte horizontales typiques du nid d'abeille. Une cellule de reine est plus grande et orientée verticalement. Si les ouvrières sentent qu'une vieille reine s'affaiblit, elles produisent des cellules d'urgence (appelées cellules de supersédure) faites à partir de cellules contenant déjà des œufs ou de jeunes larves et qui dépassent du rayon.
Quand la reine vierge émerge, elle cherche d'autres cellules de reine et pique les reines encore dedans; Si deux reines émergent simultanément, elles combattent jusqu'à la mort. La reine affirme son contrôle sur les abeilles ouvrières en libérant une suite complexe de phéromones connue sous le nom de parfum de reine.
Après plusieurs jours d'orientation dans et autour de la ruche, la jeune reine vole à un point de rassemblement des faux bourdons - un site près d'une clairière et généralement à environ 9 m au-dessus du sol - où les mâles de différentes ruches se rassemblent. Ils détectent la présence d'une reine dans leur zone de congrégation par son odeur, la trouvent par la vue et s'accouplent avec elle dans l'air; Les mâles peuvent être induits à s'accoupler avec des reines «fictives» portant la phéromone de la reine. Une reine s'accouple plusieurs fois, et peut s'accoupler plusieurs jours d’affilée (si le temps le permet) jusqu'à ce que sa spermathèque soit pleine.
Bien que la durée de vie moyenne d'une reine dans la plupart des sous-espèces soit de trois à cinq ans, les rapports de la sous-espèce d'abeille noire germano-européenne souvent utilisé pour l'apiculture indiquent qu'une reine peut vivre jusqu'à huit ans. Parce que le magasin de sperme d'une reine est épuisé près de la fin de sa vie, elle commence à pondre plus d'œufs non fécondés; Pour cette raison, les apiculteurs remplacent souvent les reines chaque année ou une année sur deux.
Les ouvrières

La durée de vie des ouvrières varie considérablement au cours de l'année dans les régions où les hivers sont longs. Les ouvrières nées au printemps et en été travaillent dur et ne vivent que 5 à 6 semaines, mais celles nées en automne resteront à l'intérieur pendant 4 à 6 mois. En moyenne pendant l'année, environ un pour cent des abeilles ouvrières d'une colonie meurent naturellement par jour. Sauf pour la reine, toutes les ouvrières d'une colonie sont remplacées tous les quatre mois environ.
Rôles de l'ouvrière d'été
L'ouvrière d'été vit un peu moins de 6 semaines. Elle passe ses 3 premières semaines à travailler dans la ruche et ses 3 dernières à butiner à l'extérieur.
- J1 à J3 : Nettoyage de l’intérieur de la ruche. L'abeille ouvrière d'été s’occupe d'abord de la préparation des cellules pour les nouvelles pontes, le temps que ses glandes nourricières se développent
- J3 à J13 : elle devient nourricière des jeunes larves avec la gelée royale qu’elle sécrète.
- J5 à J20 : elle devient cirière avec un pic de production à J12. Ses glandes nourricières s'atrophient pendant que les glandes cirières se développent. Elle participe alors à l’agrandissement des rayons, au nettoyage.
- J6 : elle réceptionne le nectar amené par les butineuses et le transforme en miel.
- J12 à J25 : elle devient gardienne pour la protection contre les prédateurs (guêpes et frelon notamment) et les voleurs (abeilles étrangères, etc.).
- J14 à J19 : elle devient ventileuse pour la régulation thermique de la ruche.
- J16 : la jeune abeille procède à son 1er vol d’orientation autour de la ruche.
- J20 à J40 : elle devient butineuse et parcourt la campagne dans un rayon de deux à cinq kilomètres afin d’approvisionner la ruche en nectar, miellat, pollen, propolis ou en eau. Après quoi, sa vie s’achève. En général, une ouvrière meurt pendant un dernier voyage de butinage, de portage d'eau (dévolu aux plus anciennes), ou pendant qu'elle dort.
Rôles de l'ouvrière d'hiver
À la fin de l'été, au début de l'automne naissent des ouvrières d'hiver, qui vont vivre de cinq à six mois. Leur corps est plus riche en acide gras. Elles ont à protéger la reine, à maintenir la colonie, qui passe l’hiver à une température de 25 à 30 °C au cœur[réf. nécessaire], puis, dès le mois de février, à préparer l'arrivée des nouvelles générations.
Bien que les abeilles ouvrières soient généralement des femelles infertiles, quand certaines sous-espèces sont stressées, elles peuvent pondre des œufs fertiles. Comme les ouvrières ne sont pas pleinement développées sexuellement, elles ne s'accouplent pas avec les faux bourdons. Les œufs des ouvrières, non fécondés, sont donc haploïdes (n'ayant que la contribution génétique de leur mère), et ces œufs haploïdes se développent toujours en faux bourdons.
Les faux bourdons


Les faux bourdons sont les abeilles mâles. Ils sont de taille supérieure aux ouvrières, car produits de la ponte (par la reine) d'œufs non fécondés dans de plus grands alvéoles. Ils naissent au printemps, quand les ruches sont actives et que de nouvelles reines non fécondées peuvent être présentes dans la ruche ou dans les environs. Ils sont dépourvus de dard. Ayant de plus gros yeux que les ouvrières, ils sont facilement reconnaissables. Ils n'ont pas de corbeille à pollen, et leur langue ne leur permet pas de recueillir le nectar des fleurs, mais ils butinent pour se nourrir. Dans la ruche, ils contribuent au maintien de la température et à la répartition de la nourriture[réf. nécessaire].
Leur développement dure 24 jours de l'œuf à l'insecte parfait, dans des conditions optimales. Même s'ils commencent à voler dès le 7e jour de leur vie d'insecte parfait, ils n'atteignent leur maturité sexuelle qu'après leur 12e jour.
Leur rôle principal est la fécondation des jeunes reines. Un faux bourdon peut produire 1,25 microlitre de sperme. Ils sont attirés par la substance chimique acide ceto-9-décene-2-oïque sécrétée par les glandes mandibulaires des reines.
À la suite de la fécondation en vol, le mâle meurt, car son abdomen est arraché pendant l'acte de reproduction. Les faux bourdons de différentes ruches du voisinage se rassemblent chaque année aux mêmes endroits, ayant des conditions thermiques spécifiques, en général dans un rayon de 4 kilomètres de leur ruche initiale. Il en résulte un brassage génétique.
En favorisant le mélange de différentes colonies, les faux bourdons sont aussi des vecteurs pour les maladies et les parasites tels que le varroa.
Leur durée de vie est d'environ deux mois, mais les abeilles peuvent consommer le couvain de faux bourdons en cas de disette. Ils sont chassés des ruches avant l'hiver, parfois dès les premiers refroidissements nocturnes d'août[7].
En cas de perte de la reine en fin de saison chaude, la ruche devient orpheline. Si aucun jeune couvain ne produit une reine, la ruche devient bourdonneuse. Des ouvrières pondent et élèvent des œufs qui, étant non fécondés, donneront naissance à des faux bourdons. Lorsque la dernière ouvrière meurt, la population uniquement constituée de faux bourdons est condamnée à mourir.
Comportement
Alimentation
Les abeilles adultes se nourrissent principalement de pollen et de miel élaboré à partir de nectar. Le miel est leur réserve de nourriture pour l'hiver. La butineuse peut aussi consommer directement le nectar prélevé sur les fleurs comme « carburant » pour retourner à la ruche.
Le miel constitue un apport énergétique (glucides). Le pollen est lui principalement constitué de protéines et de lipides. Le pollen est donc indispensable à l'ouvrière notamment dans les premiers jours de sa vie pour terminer le développement de son organisme. Les nourrices aussi utilisent de grandes quantités de pollen pour fabriquer les protéines de la gelée royale. La consommation mixte de miel et de pollen est indispensable à l'abeille. Si elle peut supporter des carences passagères de pollen, elle doit disposer de miel (ou de sucre) en permanence.
Les ouvrières adultes échangent également de la nourriture par trophallaxie. Les reines peuvent s'alimenter elles-mêmes de miel (ou de candi donné par l'apiculteur), dans la colonie, mais pendant toute leur vie, elles sont le plus souvent nourries par les ouvrières, d'un mélange de gelée royale et de miel. La reine est alimentée au pro rata du volume de sa ponte.
Pendant les trois premiers jours de leur stade larvaire, les ouvrières et les reines sont nourries de gelée royale. Les ouvrières passent ensuite à un régime de pollen et de nectar (ou de miel dilué), tandis que les reines continueront de recevoir de la gelée royale (ce qui aide les larves grosses et sexuellement développées à atteindre le stade pupal puis adulte plus rapidement). N'importe quelle larve peut devenir une reine si elle est nourrie exclusivement de gelée royale. On a longtemps pensé que la composition de la gelée royale était ce qui permettait à certaines larves de devenir des reines mais c'est en fait l'absence d'alimentation en pollen qui permet le développement supérieur des organes génitaux[8].
La colonie se développera d'autant mieux qu'elle sera située dans un endroit riche en plantes mellifères dans une zone de 3 km à la ronde de la ruche. En effet, une colonie d’abeilles a besoin, chaque saison, de plus ou moins 140 kg de nectar et 20 kg de pollen.
Si les ressources en pollen et en nectar sont nombreuses au printemps, elles sont parfois plus rares en été et en automne. Cette pénurie saisonnière peut déclencher des pillages des colonies les plus fortes sur les colonies les plus faibles. C’est pour éviter ces pillages que les apiculteurs fusionnent souvent deux colonies faibles pour en faire une forte.
Thermorégulation
L'abeille a besoin d'une température corporelle interne de 35 °C pour voler. Cette température est maintenue dans le nid pour développer le couvain[9]. C'est aussi la température optimale pour la création de la cire. Lorsqu'il fait froid, les abeilles se regroupent au centre du nid et forment une grappe compacte. La température à la périphérie de cette grappe varie avec la température de l'air extérieur, et la température interne de la grappe hivernale peut être aussi basse que 20 22 °C. Pour ne pas abaisser la température du couvain qui a besoin de chaleur pour bien se développer, les apiculteurs évitent d'ouvrir leurs ruches avant l'arrivée de températures régulières au-dessus de 15 °C soit généralement à partir du milieu du printemps en France métropolitaine.
Les abeilles peuvent se nourrir à une température de 30 °C en raison de mécanismes comportementaux et physiologiques pour réguler la température de leurs muscles de vol[réf. nécessaire]. En fonction de la température de l'air, les abeilles peuvent frissonner pour se réchauffer avant le vol ou s'arrêter pendant le vol pour frissonner à nouveau, ou s'arrêter pour se refroidir par évaporation. Les températures corporelles varient en fonction de la caste et de l'alimentation allouée correspondante. La température optimale de l'air pour la nourriture est de 22-25 °C. Pendant le vol, les muscles de vol relativement grands de l'abeille créent la chaleur qui doit se dissiper. L'abeille utilise un refroidissement par évaporation pour libérer de la chaleur à travers sa bouche. Dans des conditions chaudes, la chaleur du thorax est dissipée à travers la tête; L'abeille régurgite une goutte de fluide interne chaud qui réduit la température de sa tête de 10 °C. Au-dessous de 7 10 °C, les abeilles sont immobiles, et au-dessus de 38 °C leur activité ralentit. Les abeilles peuvent tolérer des températures allant jusqu'à 50 °C pendant de courtes périodes.
Au Canada, certains apiculteurs font hiverner leurs ruches dans des entrepôts ventilés (pour évacuer le CO2) à une température et un niveau d'humidité supportables pour les abeilles.
La danse des abeilles
Les abeilles possèdent entre elles un langage d'une extrême précision et découvert par Karl von Frisch, celui de la danse des abeilles.
Cette danse est exécutée dans l'obscurité par l'abeille éclaireuse revenue à la ruche pour renseigner les autres sur la distance, la direction, la quantité et la nature de nourriture.
Les autres abeilles, grâce à leurs perceptions tactiles et olfactives, perçoivent l'agitation et viennent s'agglutiner à elle pour décoder les informations contenues dans ces mouvements.
Il y a deux types de danses différentes :
- la danse en rond : si la nourriture se trouve à moins de 100 m, dans ce cas la direction n'est pas indiquée.
- la danse frétillante : si la nourriture se trouve à plus de 100 m, l'abeille transmet deux informations : la distance et la direction de la source de nourriture.
La nature de la nourriture est indiqué par l'odeur de l'abeille qui s'y est frottée.
La quantité de nourriture dépend du frétillement de l'abeille : plus elle frétille, plus la quantité est importante.
La distance qui sépare la source de nourriture de la ruche est transmise en fonction de la vitesse à laquelle l’abeille tourne. Plus la danse est rapide plus la source est proche.
La direction (angle entre la source de nourriture et l'aplomb du soleil par rapport à la position de la ruche) est transmise par l'inclinaison de la danse par rapport à la verticale. La précision est ± 3°. Quand la danse se prolonge, l'abeille danseuse corrige son angle en fonction de la course du soleil, et ce malgré l'obscurité complète, dans laquelle elle se trouve.
Le spectre visible de l'abeille s'étend dans l'ultraviolet (au moins jusqu'à une longueur d'onde de 300 nm)[10] donc les nuages cachant le soleil ne sont pas gênants pour elles.
Lors de l'essaimage, 300 à 500 ouvrières (souvent les butineuses les plus âgées et expérimentées) se dispersent afin de trouver un lieu pour installer la colonie (cavité). Lorsqu'une éclaireuse trouve un lieu propice à l'installation de la colonie, elle l'inspecte, durant le plus souvent moins d'une heure, à l’extérieur comme à l’intérieur et estime le volume de la cavité, la taille de l'entrée et son positionnement, sa hauteur, son exposition au vent et au soleil. Elle rentre alors à l'essaim et fait partager sa découverte grâce à une danse frétillante à la surface de l'essaim. D'autres abeilles vont alors visiter les lieux et si elles en sont satisfaites, elles le dansent à leur tour. Certains sites sont peu à peu délaissés tandis que d'autres obtiennent de plus en plus de « supporters ». L'enthousiasme des danseuses est décisif pour le choix de l'habitat, plus le site est proche des critères d’excellence plus la danse est longue et intense. Une éclaireuse qui revient de la découverte d'une habitation idéale fait en moyenne quatre-vingt dix tours d'une danse qui dure environ 135 secondes ; tandis qu'une autre ayant trouvé un site possible mais pas idéal dansera pendant 45 secondes et effectuera 30 tours. La danse d'une éclaireuse peut parfois être interrompu par une de ses sœurs qui la heurte à la tête en émettant un son bref jusqu'à ce que l'éclaireuse cesse de danser. Le lieu le plus adapté finit par obtenir l'adhésion de toutes et l'essaim s'envole vers sa nouvelle demeure.
Phéromones
Les phéromones (substances impliquées dans la communication chimique) sont essentielles à la survie des abeilles mellifères. Les abeilles s'appuient sur les phéromones pour pratiquement tous les comportements, y compris l'accouplement, l'alarme, la défense, l'orientation, la reconnaissance des familles et des colonies, la production alimentaire et l'intégration des activités des colonies.
Agressivité
Les abeilles ouvrières ont un ovipositeur modifié, un dard avec lequel elles défendent la ruche. À la différence des abeilles de tout autre genre et des reines de leur espèce, cet aiguillon ressemble à un harpon. Contrairement à la croyance populaire, une abeille ne meurt pas toujours après avoir piqué; Cette idée fausse est basée sur le fait qu'une abeille mourra habituellement après avoir piqué un humain ou un autre mammifère à peau épaisse car son dard et son sac de venin sont arrachés quand elles les piquent. Mais pour d'autres cibles plus petites et à la peau plus souple (par exemple lors de combats entre abeilles), le dard se désharponne, de sorte que l'abeille peut alors piquer puis repartir indemne (ou piquer à nouveau).
Conflit entre reine et ouvrières
Histoire
Les premiers ancêtres d'Apis mellifera sont originaires d'Afrique d'où ils ont essaimé deux fois : une première vague de migration vers l’ouest en Europe, une seconde (voire plusieurs) vers l'est, l’Asie et l’Europe orientale[11].
Au cours de l'histoire, l'abeille a évolué grâce à une forte diversité génétique. En effet, la reine de cette espèce s'unit au cours du vol nuptial à une ou deux douzaines de faux bourdons (phénomène de polyandrie), puis pond des ovules non fécondés donnant des mâles, et des ovules fécondés donnant des ouvrières qui forment une ou deux douzaines de groupes de demi-sœurs. De plus, il y a vingt fois plus de recombinaisons génétiques lors de la formation des ovules de cette espèce que dans ceux de la femme[12].
Cette diversité donne un avantage adaptatif aux colonies de cette abeille : émergence de processus coopératifs plus efficaces, d'une plus grande productivité[13].
En 2006, le consortium international Honey Bee Genome Sequencing Consortium (en) publie le premier séquençage complet du génome d'Apis mellifera, cette abeille devenant ainsi le quatrième insecte à être séquencé après la drosophile en 2000, l'anophèle en 2002 et le ver à soie en 2004[14].
Espèce menacée
Maladies et parasites
- Apocephalus borealis est un diptère qui dépose ses œufs dans l'abdomen des abeilles qui une fois infectées abandonnent la ruche puis meurent.
- Crithidia mellificae est un trypanosoma unicellulaire de l'abeille qui n'entraine pas de mortalité spécifique pour l'abeille.
- Tropilaelaps clareae est un acarien hématophage parasite externe du couvain operculé. Il peut causer la mort de la moitié du couvain. L'infestation est une MRC (maladie réputée contagieuse).
- Ascophaera apis est un champignon hétérothallique agent de la mycose du couvain ou ascosphérose (couvain plâtré, calcifié).
- Acarapis woodi est un acarien hématophage qui s'introduit dans le système respiratoire des abeilles et provoque une acariose des trachées.
- Aethina tumida est un coléoptère ravageur pour les colonies en Europe. L'infestation est une MRC (maladie réputée contagieuse).
- Braula cœca appelé poux de l'abeille est une mouche aptère qui n'entraine pas de mortalité spécifique pour les abeilles. Il a presque disparu en France.
- Varroa destructor est un acarien, le principal parasite de l'abeille, qui se nourrit de ses tissus adipeux[15].
- La maladie noire ou paralysie chronique des abeilles (CBPV, chronic bee paralysis virus), une maladie virale incurable qui atteint de très nombreuses colonies.
- Nosema apis est un champignon unicellulaire qui envahit le ventricule de l'abeille et provoque une dysenterie ou paralysie. Il peut être un facteur du syndrome d'effondrement.
- Nosema ceranae est un champignon qui provoque une nosémose.
- le Virus israélien de la paralysie aiguë IAPV (Israeli acute paralysis virus) infecte les abeilles et entraine une paralysie.
- *Les larves des diptères de la famille des Conopidae sont des endoparasites obligatoires d'hyménoptères. Pour se faire, les femelles coursent et agressent violemment leur hôte afin de lui déposer un œuf sur le corps. Se développant à l'intérieur de son abdomen, la larve se nourri des muscles du thorax. En Europe, Apis mellifera subit les affres de Physocephala rufipes, P. vittata, Merziella longirostris et Zodion cinereum[16]. Vespa velutina nigrithorax, quant à lui, est logé à la même enseigne par l'intermédiaire de Conops vesicularis[17].
Effondrement des colonies d'abeilles
- Les abeilles subissent depuis le début du XXIe siècle des pertes importantes dans toutes les régions du monde.
Les scientifiques et les apiculteurs de l'UNAF estiment que les produits phytosanitaires utilisés par l'agriculture intensive affaiblissent voire tuent les abeilles[18]. Ces pesticides pourraient empêcher les abeilles de lutter efficacement contre les maladies et parasites des ruches (ex: nosémose) et parasitoses (ex: Varroa). La durée de vie des reines est passée de quatre ans il y a quelques décennies à moins de deux ans maintenant (probablement à cause de la nourriture polluée)[réf. nécessaire]. - La DGAL estime pour sa part, d'après les résultats de ses enquêtes annuelles, que les principaux facteurs de la mortalité des abeilles en France sont, par ordre d'importance, les pathologies (dont majoritairement le varroa destructor) dans au moins 40% des cas, suivies des pratiques apicoles (négligences, mauvais traitement des pathologies, etc.), dans 14% des cas, et du manque de ressources alimentaires conduisant à la famine des ruches. Enfin, les pratiques phytosanitaires ne seraient responsables que dans 4% des cas. Parmi les principaux produits incriminés figurent le Spinosad, insecticide utilisé en agriculture biologique, mais aussi des cires à usage agricole, notamment en traitement anti-varroa à l'intérieur des ruches ou comme insecticide pour contrôler des ravageurs du colza[19].
- Le nombre d'abeille domestique a cependant augmenté dans le monde de 30% depuis 2000 et de 45% en un demi siècle[20].
- La disparition des abeilles met de nombreux écosystèmes et agrosystèmes en danger car l'abeille est un vecteur essentiel dans le processus de pollinisation (dont agricole pour par exemple la production de luzerne, fruit, légume, etc. qui augmente de 20 % environ en présence d'abeilles ; sans abeille les amandiers, poiriers, fraisiers, etc. ne produisent presque plus).
- L’hybridation par des importations d’abeilles d’autres sous-espèces, moins adaptées à notre environnement, a pu rendre les abeilles locales plus fragiles[réf. nécessaire].
Coexistence entre abeilles semi-domestiques et sauvages
Des colonies sont souvent installées ou déplacées de milieux semi-naturels vers des milieux naturels, voire introduites dans des aires naturelles protégées ou des espaces naturels sensibles riches en fleurs, ce qui peut poser des problèmes de cohabitation avec les abeilles sauvages (dont près de 1000 espèces étaient en 1995 recensées en France, 380 espèces en Belgique, 587 en Suisse et 274 au Luxembourg[21]), rappelle[22] l’Observatoire des Abeilles [23]. Cette cohabitation forcée entraîne trois types de risques :
- une compétition avec les abeilles sauvages (pour la ressource en pollen et nectar) ; Ces dernières sont défavorisées par l'apiculture. Or les données scientifiques montrent que les apidés sauvages autochtones sont souvent de meilleurs pollinisatrices des fleurs sauvages[24] et cultivées [25],[26],[27],[28].
- des transmissions de maladies contagieuses (et/ou de parasites), de l'abeille mellifère vers les autres espèces (ou inversement) ;
- « une modification des patrons de flux de pollen entre plantes pouvant entraîner une perturbation de la reproduction des plantes et une modification de la composition des communautés végétales », au détriment de la biodiversité et des services écosystémiques ;
- et parfois (localement) une exposition accrue à des prédateurs (guêpiers...).
Cette coexistence n'est toutefois pas nouvelle. L'abeille mellifère est domestiquée par l'homme depuis des milliers d'années sans conséquences négatives de grande envergure pour la faune et la flore sauvage. Ces risques sont par ailleurs difficiles à évaluer car ils dépendent fortement du nombre de colonies, de leurs localisations et des périodes considérées. On notera par exemple que dans certaines régions du monde, en Afrique et en Asie, l'abeille mellifère est indigène et présente à l'état sauvage.[29]
Races et sélection génétique
L'abeille à miel a un génome différent selon le sexe. La femelle (la reine et les ouvrières) est diploïde. Son génome est composé de 2n=32 chromosomes[30]. Le mâle (le faux bourdon), issu d'un œuf non fécondé, n'a lui que 16 chromosomes et est donc haploïde. Les mâles produits à partir d'une reine ne contiennent que ses gènes. Ceci peut être utilisé avantageusement dans les programmes de sélection. Même les reines fécondées naturellement peuvent être utilisées pour produire des mâles purs de la lignée de la reine.
Chaque abeille est unique en raison de la recombinaison très importante des chromosomes de la reine lors de la formation des œufs (méiose). Il a été constaté qu'il y a beaucoup plus de recombinaisons ou de «croisements» de chromosomes chez les abeilles domestiques que dans tout autre animal étudié jusqu'à présent. La recombinaison est une source de variation, et la variation est la matière première de l'adaptation. C'est peut-être ce qui a permis à l'abeille de s'adapter à presque toutes les régions de la Terre[31].
Les sélectionneurs du monde entier tentent d'améliorer la génétique de l'abeille pour obtenir des variétés plus productives en miel et plus résistantes aux maladies et aux différents problèmes environnementaux. La majorité des variétés sont nées naturellement en s'adaptant à leur milieu mais il existe aussi des variétés « synthétiques » comme la Buckfast qui ont été créées par l'Homme par croisement multiples de variétés naturelles.
Les qualités et les caractéristiques désirées pour l'élevage peuvent être divisées en trois groupes : primaire, secondaire et tertiaire, ceux qui ont une incidence sur la gestion.
Critères primaires
Les qualités primaires sont les qualités essentielles à une production maximale de miel.
- Fécondité - maintien d'au moins 9 cadres de couvaison de mai à juillet
- Zèle fourrager - une capacité illimitée pour le travail de la nourriture, la consanguinité étroite pour intensifier cette qualité peut être contre-productive.
- Résistance aux maladies
- Peu enclin à l'essaimage
Critères secondaires
- Longévité
- Puissance des ailes
- Sens de l'odorat
- Caractéristiques défensives
- Rusticité et aptitude à hiverner sans consommer trop de miel
- Développement printanier
- Instinct d'auto-approvisionnement
- Arrangement des rayons de miel
- Production de cire et construction des rayons
- Collecte de pollen
- Longueur de la langue (pour pouvoir polliniser certaines fleurs plus profondes)
Critères tertiaires
- Bon tempérament, comportement calme
- Désinclination à la propolisation
- Pas de collage de rayons
- Propreté, hygiène sensible aux varroas (VSH)
- Operculation du miel
- Sens de l'orientation
Abeille et qualité de l'air
Abeilles contribuant à améliorer la qualité de l'air
En collectant de grandes quantités de pollen, les abeilles diminuent la quantité de pollen allergène (ou devenant allergène en vieillissant et/ou en se dégradant).
Ceci a notamment été montré au début des années 2000 dans les serres de culture du poivron aux Pays-Bas (où environ 8 000 ouvriers agricoles le cultivent dans 10 000 hectares de serre). La fleur du poivron produit un pollen irritant auquel les travailleurs des serres sont exposés quand ils les cueillent. Dans les serres classiques, 1/3 de ces travailleurs finissent par devenir allergiques à ce pollen. Mais on a montré qu'introduire une colonie d'abeille domestique (ruche) dans la serre suffit à faire chuter le taux d'allergie et sa gravité[32] ; Dans les serres où les abeilles étaient présentes, le pollen était récolté par les abeilles sur les anthères au fur et à mesure de sa production, et les symptômes d'allergies diminuaient ou disparaissaient chez les travailleurs. L'étude n'est cependant pas allé jusqu'à calculer le nombre minimal ou optimal d'abeilles par serre.
Abeilles victimes de la pollution de l'air
Tous les apidés (ou leurs larves) sont victimes de divers insecticides aéroportés.
Des études les ont proposé et utilisé comme bioindicateur pour certaines retombées ou émissions radioactives[réf. nécessaire]
Elles sont aussi exposées à des métalloïdes et métaux lourds toxiques, dont le plomb. Les taux d'éléments traces métalliques (As, Mn, Pb et Zn) et las compositions isotopiques du Pb ont été relevés sur des échantillons d'abeilles, de cire et de miel provenant de sols urbains (Sydney) et jouxtant un parc national (Broken Hill) en Australie. Dans tous les cas le métal le plus contaminant était le Plomb (provenant du sol et de la poussière) trouvé à des taux correspondants chez les abeilles mellifères, leur cire et leur miel[33]. Le taux de plomb du sol était le meilleur facteur prédictif de la contamination des abeilles et de leurs produits. Les analyses isotopiques du plomb (206Pb/207Pb et 208Pb/207Pb) ont en outre montré que :
- le plomb trouvé dans les abeilles et dans leurs production était isotopiquement presque identiques (95-98 %) au plomb de leur environnement[33] ;
- les abeilles mellifères rurales étaient contaminées par un plomb dont l'origine était géogénique et locale, alors que les abeilles de la région métropolitaine de Sydney étaient plus contaminées ; et contaminées par un plomb dont la signature isotopique est comparable à celle des aérosols mesurés pendant la période d'utilisation de l'essence au plomb[33].
Ceci confirme que :
- l'abeille est un bio indicateur pertinent pour l'évaluation de la contamination historique ou contemporaine de l'air par le plomb, que ce plomb soit naturel et/ou anthropique (c'est-à-dire « environnementalement hérité ») ; à condition que les sources soient bien documentées[33] ;
- bien après l'interdiction du plomb dans l'essence, ce plomb « hérité », non-dégradable et séquelle de pollutions anciennes continue à être remobilisées via les envols de poussière, contaminant ainsi les systèmes alimentaires et écologiques[33].
Notes et références
- (en-US) « What's missing from claims that neonicotinoids are killing bees, birds and fish? », sur Genetic Literacy Project, (consulté le )
- (en) Irati Miguel, Mikel Iriondo, Lionel Garnery, Walter S. Sheppard et Andone Estonb, « Gene flow within the M evolutionary lineage of Apis mellifera : role of the Pyrenees, isolation by distance and post-glacial re-colonization routes in western Europe », Apidologie, vol. 38, , p. 141-155 (DOI 10.1051/apido:2007007).
- Quelle abeille choisir pour son rucher ?
- « Bienvenue sur le site ApiClass », sur apiclass, Muséum national d'histoire naturelle, France, site d'identification des variétés d'abeilles
- « Les critères de différenciation des races d'abeilles », sur apihappy.fr
- « Les races d’abeilles », sur catoire-fantasque.be,
- http://www.apiculture.com/.
- Gwen Pearson, « Royal Jelly Isn't What Makes a Queen Bee a Queen Bee », Wired, (ISSN 1059-1028, lire en ligne, consulté le )
- Comment les abeilles contrôlent-elles la température de la ruche?
- (en) Adriana D. Briscoe et Lars Chittka, « The evolution of color vision in insects », Annual Review of Entomology, vol. 46, , p. 471-510 (ISSN 0066-4170, résumé).
- (en) E. Pennisi, « GENETICS : Honey Bee Genome Illuminates Insect Evolution and Social Behavior », Science, vol. 314, no 5799, , p. 578-579 (DOI 10.1126/science.314.5799.578).
- (en) G. J. Hunt et R. E. Page-Jr, « Linkage Map of the Honey Bee, Apis Mellifera, based on Rapd Markers », Genetics, vol. 139, no 3, , p. 1371–1382 (lire en ligne).
- (en) Mattila HR, Seeley TD, « Genetic diversity in honey bee colonies enhances productivity and fitness », Science, vol. 217, no 3, , p. 362–364 (DOI 10.1126/science.1143046).
- (en) George M. Weinstock, Gene E. Robinson et col, « Insights into social insects from the genome of the honeybee Apis mellifera », Nature, vol. 443, no 7114, , p. 931–949 (DOI 10.1038/nature05260).
- (en) Samuel D. Ramsey, Ronald Ochoa, Gary Bauchan, Connor Gulbronson, Joseph D. Mowery, Allen Cohen, David Lim, Judith Joklik, Joseph M. Cicero, James D. Ellis, David Hawthorne et ennis vanEngelsdorp, « Varroa destructor feeds primarily on honey bee fat body tissue and not hemolymph », PNAS, vol. 116, no 5, , p. 1792-1801 (lire en ligne, consulté le ).
- (en) Jens-Hermann Stuke, Conopidae (Diptera), World Catalogue of Insects, Volume: 15, Brill, 2017, (ISBN 978-90-04-27183-8)
- (en) Darrouzet E., Gévar J., Dupont S., « A scientific note about a parasitoid that can parasitize the yellow-legged hornet, Vespa velutina nigrithorax, in Europe. », Apidologie, vol. 46, , p. 130-132 (DOI 10.1007/s13592-014-0297-y)
- Dossier de presse compilé par l'Union nationale de l'apiculture française, 2010.
- Vincent Jay, « Bilan des suivis des mortalités d'abeilles par la DGAL (Ministère de l'Agriculture) : Un communiqué du Réseau Biodiversité pour les Abeilles », Média Terre, (lire en ligne, consulté le ).
- (en) Jonathan Kay, « The Insect Apocalypse That Never Was »,
- Rasmont P, Ebmer AW, Banazsak J & Van der Zanden G, 1995. Hymenoptera Apoidea Gallica. Liste taxonomique des abeilles de France, de Belgique, de Suisse et du Grand-Duché de Luxembourg. Bulletin de la Société Entomologique de France (hors série)100 : 1–98.
- Nicolas J. Vereecken, Eric Dufrêne & Michel Aubert (2015) Sur la coexistence entre l'abeille domestique et les abeilles sauvages ; Rapport de synthèse sur les risques liés à l’introduction de ruches de l’abeille domestique (Apis mellifera) vis-à-vis des abeilles sauvages et de la flore ; Observatoire des abeilles, 29 mai 2015, PDF, 21 pp, accessible sur www.oabeilles.net.
- l’Observatoire des Abeilles (OA, www.oabeilles.net) est une ONG (association loi 1901) ayant pour objet l'étude de l’écologie et de la conservation des abeilles sauvages dans leurs habitats naturels.
- Larsson M (2005) Higher pollinator effectiveness by specialist than generalist flower-visitors of unspecialized Knautia arvensis (Dipsacaceae)' ; Oecologia. décembre 2005 ; 146(3):394-403. Epub 2005 Oct 27. (résumé).
- Banda HJ & Paxton RJ, (1991) Pollination of greenhouse tomatoes by bees. Acta Horticulturae 288: 194-198.
- Willmer PG, Bataw AAM & Hughes JP, 1994. The superiority of bumblebees to honeybees as pollinators – Insect visits to raspberry flowers. Ecological Entomology 19: 271-284.
- Buchmann SL & Nabhan GP, 1996. The forgotten pollinators. Island Press, Shearwater books, Washington DC.
- Richards KW, 1996. Comparative efficacy of bee species for pollination of legume seed crops. In: The conservation of bees (A. Matheson, S. Buchmann, C. O’Toole, P. Westrich & I. H. Williams, Eds.), p. 81-103, Academic Press, London.
- « Apiculture et moyens d'existence durables », sur www.fao.org (consulté le )
- Video : Why genetics are important to beekeeper ?
- Genetic Aspects of Queen Production.
- S. van der Steen, T. Blacquière, N. de Jong, H. de Groot, honey bees as an aid in improving labour conditions in sweet bell pepper greenhouses : Reduction of pollen allergy ; ISHS Acta Horticulturae 639: XXVI International Horticultural Congress: Expanding Roles for Horticulture in Improving Human Well-Being and Life Quality, 2004 (Résumé, en anglais).
- Zhou, X., Taylor, M. P., Davies, P. J., & Prasad, S. (2018). Identifying sources of environmental contamination in European honey bees (Apis mellifera) using trace elements and lead isotopic compositions. Environmental science & technology, 52(3), 991-1001.
Voir aussi
Articles connexes
Références taxonomique
- Ressources relatives au vivant :
- Fauna Europaea
- Nederlands Soortenregister
- TAXREF (INPN)
- (en) ARKive
- (cs + en) BioLib
- (en) BugGuide
- (en) Catalogue of Life in Taiwan
- (sv) Dyntaxa
- (en) EPPO Global Database
- (en) EU-nomen
- (en) European Nature Information System
- (en) Fossilworks
- (en) Global Biodiversity Information Facility
- (en) iNaturalist
- (en) Interim Register of Marine and Nonmarine Genera
- (cs) Nálezová databáze ochrany přírody
- (en) NBN Atlas
- (en) New Zealand Organisms Register
- (en) Système d'information taxonomique intégré
- (en) ZooBank
- (fr+en) Référence ITIS : Apis mellifera Linnaeus
- (en) Référence Hymenoptera Online Database : Apis mellifera
Liens externes
- (fr) Site de l'association belge d'apiculture CARI
- (fr) Conservation et élevage de l'abeille noire
- (fr) Union Nationale des Apiculteurs Français
 Portail de l’agriculture et l’agronomie
Portail de l’agriculture et l’agronomie  Portail de l’entomologie
Portail de l’entomologie