Acarinarium
Un acarinarium (pluriel : acarinaria ou acarinariums), est une structure anatomique spécialisée qui facilite la rétention d'acariens dans le corps d'un organisme, particulièrement au sein du sous-ordre hyménoptère Apocrita (abeilles et guêpes dans leur sens très large).
La nature exacte de leur relation n’est pas entièrement comprise. La présence d'un acarinarium concerne souvent uniquement les femelles de l'espèce hôte et est utilisée comme critère de détermination[1]. Ce terme a été créé en 1920 par Walter Karl Johann Roepke (1882-1961), entomologiste allemand notamment spécialisé dans les coléoptères et lépidoptères indomalais et javanais[2].
Ectosymbiose



Des fossiles d'abeilles du genre éteint Oligochlorade (Halictidae) muni d'un acarinarium ont été trouvés dans un dépôt d'ambre dominicain[3]. L'acarinarium a évolué afin d'améliorer la symbiose entre les acariens et leur hôte. Il y a de nombreux cas où les Acariens sont phorétiques (c'est-à-dire qu'ils vivent accrochés à leur hôte) sur des organismes qui bénéficient de leur présences, mais les cas où le corps de l'hôte s'est adapté afin d’accueillir en son sein les acariens sont beaucoup plus rares. Il s'agit d'ectosymbiose.
Les nids d'Hyménoptères représentent des micro-biotopes où de multiples proies, prédateurs ou fossoyeurs se développent. Chez les Apocrita, il semble que les acariens favorisé par l'acarinarium se nourrissent de champignons dans les nids des hôtes (les éloignant ainsi de leurs larves ou de leurs provisions) ou éventuellement d'autres parasites ou acariens dont la présence dans le nid est préjudiciable aux hôtes ou encore de déchets tel que le pollen. Dans la plupart des cas, seules les femelles possèdent un acarinarium car ce sont elles qui construisent et fournissent les nids[1],[4]. Plusieurs espèces d'acariens peuvent être présentes en même temps dans l'acarinarium, et en adjonction avec des acariens phorétiques. Dans d'autres cas, ils peuvent être monospécifiques[4]. Étant donné qu'il y a plusieurs tailles d'Acariens, la taille de l'acarinarium influe sur l'espèce accueillie[5].
La nature exacte de leur symbiose n’est pas entièrement comprise, leur relation pourrait relever du commensalisme[1],[4] ou du mutualisme[1],[4]. D'autres, suggèrent qu'il s'agit d'une réponse au parasitisme. Les premiers nids cloisonnés des Apidées étant plus infestés que les autres, l'acarinarium servirait alors de réservoir piège réduisant ainsi les risques de dispersion et leur effet sur les autres abeilles de la couvée[6]. Enfin, un « mutualisme conditionné » a pu être étudié chez une Guêpe potière Allodynerus delphinalis. L’acarien symbiotique Ensliniella parasitica (Sarcoptiformes) présent dans leur acarinarium ingère l’hémolymphe des guêpes juvéniles, mais les protège également d’un ennemi naturel, Melittobia acasta (Eulophidae). Or, lorsque ce dernier est peu présent, le comportement de l'Acarien dérive vers le parasitisme en diminuant le développement des mâles et en augmentant la mortalité des juvéniles[7].
Différentes formes d’acarinarium
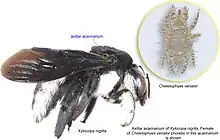
Différentes formes d’acarinarium ont évolué au sein de différentes lignées d’Abeilles charpentières, dans les sous-genres eurasiatiques, Koptortosoma et Mesotrichia. Dans certains cas, une grande cavité abdominale lui est consacrée, quand d'autres espèces lui dédient également le premier tergite du métasome[1]. D'autres encore, présentent un acaridarium axillaire.
Le genre Thectochlora de la famille des Halictidae, se caractérise par un pinceau de poils dense en face de la face antérieure du premier tergite du métasome (emplacement identique aux Abeilles charpentières), ce qui crée un espace semblable à une pochette dans lequel les acariens sont transportés[1].
Dans le sous-genre Lasioglossum, autres Halictidae, de nombreuses espèces n'ont plus de soies centrales sur la face antérieure du premier tergite du métasome, formant ainsi une zone centrale glabre autour de laquelle se trouve une frange de poils. Les espèces sans cette modification ont rarement, voire jamais, d'acariens dans cette zone, tandis que celles qui ont cette région glabre ne manquent presque jamais d'acariens. Ainsi, malgré sa nature simple, il semble bien fonctionner comme un acarinarium. Dans certains cas, des espèces sœurs peuvent différer par ce trait, l'une ayant un acarinarium et l'autre non, ce qui suggère que ce détail peut être perdu et éventuellement retrouvé au cours de l'évolution[1].
Chez les Guêpes potières du genre Parancistrocerus, la base du deuxième tergite du métasome est concave et recouverte par la lèvre postérieure du premier tergite métasomal. Les acariens sont ainsi véhiculés dans la concavité. Cette structure semble la même chez les mâles et les femelles, bien qu'il soit difficile d'évaluer si les deux sexes sont porteurs d'acariens à une fréquence égale[8].
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Acarinarium » (voir la liste des auteurs).
- (en) Charles Duncan Michener, 2000, The Bees of the World, Johns Hopkins University Press (ISBN 0801861330)
- (nl) Walter K.J. Roepke, « Verslag van de drie-en-vijftigste wintervergadering der Nederlandsche Entomologisch Vereeninging » [« Rapport du 53e congrès hivernal de la Société entomologique néerlandaise »], Tijdschrift voor Entomologie, vol. 63, , p. 11-18.
- (en) Michael S. Engel & Molly G. Rightmyer, 2000, A new augochlorine bee species in Tertiary amber from the Dominican Republic (Hymenoptera: Halictidae), Apidologie, volume 31, pages 431–436, Doi
- (en) Barry M. OConnor, 1993, The mite community associated with Xylocopa latipes (Hymenoptera: Anthophoridae: Xylocopinae) with description of a new type of acarinarium, International Journal of Acarology Volume 19, Issue 2, Pages 159-166, Doi
- (en) Kimiko Okabe & Shun’ichi Makino, 2002, Phoretic mite fauna on the large carpenter bee Xylocopa appendiculata circumvolans (Hymenoptera: Apidae) with descriptions of its acarinaria on both sexes, Journal of the Acarological Society of Japan, volume 11, issue 2, pages 73–84, Doi
- (en) Pavel B. Klimov, S. Bradleigh Vinson and Barry M. OConnor, 2007, Acarinaria in associations of apid bees (Hymenoptera) and chaetodactylid mites (Acari). Invertebrate Systematics Volume 21, Issue 2, pages 109–136 Doi
- (en) K. Okabe, S. Makino, 2010, Conditional Mutualism Between Allodynerus delphinalis (Hymenoptera: Vespidae) and Ensliniella parasitica (Astigmata: Winterschmidtiidae) May Determine Maximum Parasitic Mite Infestation, Environmental Entomology, Volume 39, Issue 2, Pages 424–429, Doi
- (en) Roberto Boesi, Carlo Polidori, Jose Tormo, Stefania Bevacqua, Josep Daniel Asis, Francesco Andriett, 2005, Trap-nesting Ancistrocerus sikhimensis (Hymenoptera:Eumenidae) in Nepal : nest structure and associates (Hymenoptera: Chrysididae; Acarina:Saproglyphidae), Florida Entomologist, volume 88, issue 2, pages 135–140, doi, Pdf
Liens externes
- (en) Barry O'Connor, « North-American Bee-associated Mites, Potentiel Threats on Native and Introduced Pollinisators », Museum of Zoology at the University of Michigan.
 Portail de l’entomologie
Portail de l’entomologie  Portail de l’arachnologie
Portail de l’arachnologie