Équation de Michaelis-Menten
L’équation de Michaelis-Menten (ou de Michaelis-Menten-Henri) permet de décrire la cinétique d'une réaction catalysée par une enzyme agissant sur un substrat unique pour donner irréversiblement un produit. Elle relie la vitesse stationnaire initiale de la réaction à la concentration initiale en substrat et à des paramètres caractéristiques de l'enzyme.
L'équation décrit un comportement cinétique très classique, observé avec de nombreuses enzymes, mais elle ne permet pas de rendre compte de certains comportements complexes, comme ceux résultant de l'existence de plusieurs sites actifs qui interagissent (voir l'article allostérie pour plus de détails), ni de comprendre les propriétés cinétiques d'enzymes qui catalysent une réaction faisant intervenir plusieurs substrats. L'équation n'est pas valable si la réaction est catalysée de façon réversible et que le produit de la réaction est présent (voir l'article Equation de Haldane pour plus de détails).
Formule et représentation

Selon le modèle de Michaelis et Menten, l'équation décrivant la vitesse initiale stationnaire d'une réaction enzymatique est la suivante :

Avec :
- : vitesse initiale (c’est-à-dire en absence de produit) stationnaire (indépendante du temps) de la réaction enzymatique pour une concentration initiale en substrat . Comme toutes les vitesses de réactions chimiques, elle a la dimension d'une concentration par unité de temps (par exemple en µmol/L/min)[1] ;
- : vitesse initiale maximale, notée sur la figure ci-contre, la vitesse de la réaction quand la concentration en substrat est très grande (on parle de concentration saturante), dans la même unité que (par exemple en µmol/L/min)[1] ;
- la vitesse initiale et la vitesse initiale maximale sont toutes les deux proportionnelles à la concentration en enzyme dans le milieu réactionnel ;
- : concentration initiale en substrat (en mol/L)[1] ;
- est la Constante de Michaelis de l'enzyme. C'est la valeur de pour laquelle la vitesse initiale est la moitié de la vitesse initiale maximale. Son unité est celle d'une concentration (mol/L).





L'ordre de réaction dépend des grandeurs relatives des deux termes au dénominateur.
- A basse concentration en substrat , de sorte que Dans ces conditions la vitesse initiale est une fonction linéaire de la concentration du substrat , et la réaction est du premier ordre par rapport au substrat.



- Au contraire, si la concentration initiale en substrat est élevée, avec , la vitesse initiale devient indépendante de (cinétique d'ordre zéro par rapport au substrat) et approche la vitesse maximale de façon asymptotique. Cette valeur maximale est atteinte dans des conditions où toute l'enzyme est liée au substrat à l'état stationnaire. Augmenter davantage la concentration initiale en substrat n'augmente pas la vitesse initiale, on dit que l'enzyme est "saturée" ou que la concentration en substrat est "saturante".


Démonstration de l'équation de Michaelis-Menten
Dans les présentations modernes de cette équation, on part de l'hypothèse de mécanisme ci-dessous :

avec les constantes de vitesse des différentes étapes.

L'analyse de Michaelis-Menten se fait à l'aide de deux hypothèses simplificatrices :
- Vitesse initiale: on considère des conditions initiales, avant que le produit n'ait le temps de s'accumuler dans le test enzymatique, et avant que la concentration en substrat n'ait le temps de diminuer significativement. Cela a deux conséquences:
- Dans le calcul on peut considérer que la concentration en S est constante: .
- On peut négliger la contribution de la réaction inverse dont la vitesse est , puisque .



Le système se simplifie alors de la manière suivante :

avec


- Deuxième hypothèse simplificatrice: l'approximation des états quasi stationnaires. Si on s'intéresse à la période ou la vitesse est stationnaire (constante), cela implique que est constante, ce qu'on peut écrire sous la forme .


Pour déterminer la vitesse de la réaction, on doit donc exprimer en fonction des paramètres du modèle: les 3 constantes de vitesse, la concentration en substrat et la concentration en enzyme . et sont les deux inconnues, pour les déterminer il faut utiliser deux équations. La première vient de l'hypothèse de stationnarité. La deuxième vient d'une hypothèse de conservation.


- Utilisation de l'hypothèse de stationnarité:
La vitesse de formation du complexe est égale à . Pendant la période stationnaire, la concentration du complexe enzyme-substrat est constante, ce qui impose:


Soit

- Utilisation de l'équation de conservation:
En plus de l'équation de l'état stationnaire, on utilise une deuxième équation qui traduit la conservation de la quantité totale d'enzyme: celle ci se retrouve soit sous sa forme "libre" E soit sous sa forme liée au substrat . En notant la concentration totale en enzyme, on a donc:

Comme on cherche à exprimer , on peut réécrire cette équation sous la forme:

soit:

D'où

On trouve l'équation de Michaelis et Menten :

à condition de poser et


La loi de vitesse de Michaelis et Menten relie la vitesse initiale de la réaction à la vitesse maximale , la constante de Michaelis , et la concentration initiale en substrat .
Interprétation de la constante de Michaelis
La constante de Michaelis est la concentration en substrat pour laquelle la vitesse initiale de la réaction est égale à la moitié de la vitesse initiale maximale (en mol/L)[1]. Elle a la dimension d'une concentration (même unité que )


Si l'on fait l'hypothèse du mécanisme suivant , alors (voir la démonstration ci-dessus).


Elle correspond à l'inverse de la constante de dissociation apparente du substrat pour l'enzyme dans certaines conditions. En effet, on constate que la définition de la constante de Michaelis peut être reformulée : , en se souvenant que la définition de la constante de dissociation est : .


On voit donc bien que, dans des conditions d'équilibre (qui elles seules permettent de mesurer des constantes d'équilibre K), la constante prend en compte la valeur de la constante de dissociation de l'enzyme pour le substrat, mais qu'en conditions catalytiques (lorsque l'enzyme peut fonctionner et que ES donne E + P), on doit prendre en compte le rapport de la constante de vitesse de l'enzyme et de la constante de dissociation non-productive de ES: . Dit autrement, lorsque l'enzyme n'est pas catalytiquement active , alors . Ainsi, il y a bien un lien entre le et le mais ils ne sont pas équivalents.




Rappelons aussi que (la constante de dissociation est l'inverse de la constante d'affinité) et donc, que le , sous ces conditions ( ), peut mesurer la constante de dissociation, et indirectement l'affinité de l'enzyme pour le substrat.

Graphiquement, la variation de la vitesse en fonction de la concentration en substrat prédite par l'équation de Michaelis-Menten est une branche d'hyperbole.
Aspects historiques
L'équation est souvent attribuée à l'Allemand Leonor Michaelis et la Canadienne Maud Menten, mais elle avait été proposée sous une forme voisine par le Français Victor Henri dès 1902[2],[3],[4]. La contribution principale de Michaelis et Menten dans ce contexte est d'avoir proposé en 1913 de mesurer et interpréter des vitesses initiales de réaction[5],[6],[7].
Par ailleurs, la démonstration basée sur l'hypothèse de l'état stationnaire et présentée sur cette page est due à Briggs et Haldane (1925)[8]. La démonstration originale de Michaelis et Menten en 1913 est en fait basée sur une hypothèse un peu différente, selon laquelle la réaction de formation du complexe enzyme/substrat est un équilibre très rapide (première ligne du tableau ci-dessous).
L'année suivante Van Slyke et Cullen obtiennent une équation identique à celle de Michaelis et Menten en faisant l'hypothèse contradictoire que la fixation du substrat est irréversible[9] (ligne 2 dans le tableau ci-dessous). (L'observation que des modèles contradictoires donnent la même équation de vitesse (l'équation dite de Michaelis et Menten) est importante: elle illustre que l'accord expérimental entre des données et une loi de vitesse ne permet pas de conclure que les hypothèses du modèle sont exactes.)
La controverse se clôt lorsque Briggs et Haldane en 1925 montrent que les deux modèles sont les cas particuliers d'un modèle plus général[8], celui qui est décrit sur cette page et dans la ligne 3 du tableau ci-dessous.
En fait, n'importe quel schéma cinétique qui enchaîne de façon séquentielle des réactions de fixation/dissociation des substrat et produit et des réactions du premier ordre donne une loi de vitesse de type Michaelis et Menten, mais la signification des paramètres et dépend du modèle.
| Auteurs | Hypothèse | Mécanisme | Loi de vitesse | ||
|---|---|---|---|---|---|
| Victor Henri, puis Michaelis et Menten, 1913[6] | La fixation du substrat est réversible, très rapide et reste à l'équilibre | ||||
| Donald Van Slyke (en) et Cullen, 1914[9] | La fixation du substrat est irréversible | ||||
| Briggs et Haldane, 1925[8] | Le cas général | ||||
| Un mécanisme plus complexe qui enchaîne deux étapes du premier ordre | |||||
| L'un des mécanismes envisagé par Victor Henri[3],[4], dont on sait aujourd'hui qu'il est faux | NB: Ici, le compexe ES n'est pas un intermédiaire catalytique et est une constante de vitesse bimoléculaire. |












Détermination des constantes
Il existe plusieurs méthodes pour déterminer les paramètres de Michaelis-Menten d'une enzyme. Dans la pratique, on réalise une série de mesures de la vitesse initiale de la réaction pour différentes valeurs de la concentration de substrat [S]. Il est important de se placer dans des conditions de vitesse initiale (absence de produit P en quantité significative) pour respecter les hypothèses simplificatrices discutées ci-dessous.
Historiquement, pour déterminer numériquement les paramètres et , les enzymologistes ont eu recours à différentes formes de linéarisation de l'équation de Michaelis-Menten. Il s'agit de changements de variables qui transforment l'équation initiale en équation linéaire, qu'il est alors possible d'ajuster graphiquement ou par régression linéaire. Parmi ces linéarisations classiques, on peut citer la représentation de Lineweaver et Burk, celle de Eadie et Hofstee ou celle de Scatchard.

Aujourd'hui, ces différentes techniques ont été remplacées par la régression non linéaire, plus précise et plus robuste.
Représentation de Lineweaver-Burk
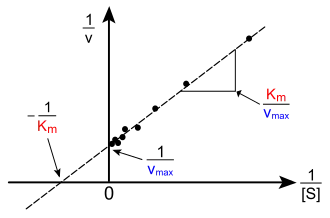
Dans cette approche, on détermine les constantes de l'enzyme et par la représentation des inverses (représentation de Lineweaver et Burk) qui est une droite de en fonction de , dont l'équation est:



La pente est alors et l'ordonnée à l'origine est ,


d'où
et


Cette représentation est une des moins précises quant à la détermination des caractéristiques de l'enzyme (Vmax, Km). En effet, les représentations de Lineweaver-Burk et Eadie-Hofstee ont une coordonnée en 1/[S]0, donc les mesures les plus précises seront concentrées dans la même région (voisines de l'axe vertical) et peu de mesures avec une erreur relativement grande existeront pour des faibles valeurs de concentration de substrat : le tracé à la règle de la "meilleure" droite sera entaché d'erreur.
Représentation de Hanes-Woolf
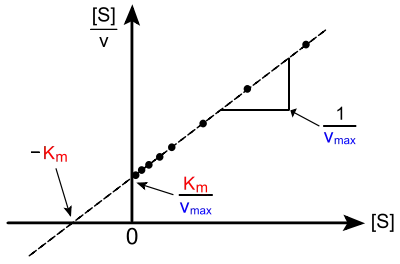
La représentation de Hanes-Woolf est considérée moins susceptible à la propagation statistique des erreurs de mesure que la représentation de Lineweaver-Burk.
La droite est d'équation :

Pour un graphique de en fonction de , la pente est et l'ordonnée à l'origine est


Représentation de Eadie-Hofstee
Le diagramme de Eadie et Hofstee indique la vitesse de réaction en fonction du rapport selon


Cette équation est obtenue à partir de l'équation de Michaelis-Menten en inversant et en multipliant par comme suit:



D'ou on peut isoler :

Et alors

Il s'ensuit qu'un graphique de en fonction de sera une ligne droite qui permet d'évaluer comme ordonnée à l'origine et comme pente.


Représentation de Eisenthal et Cornish-Bowden
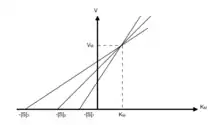
L'équation de la droite est :

Ainsi, sur le graphique de en fonction de , pour chaque valeur de correspondante, on reporte sur l'axe des abscisses la valeur de -. On obtient plusieurs droites qui se recoupent en un même point de coordonnées (ordonnée) et (abscisse).


Articles connexes
Notes et références
- Le choix des unités est arbitraire et doit rester homogène
- (en) Athel Cornish-Bowden, Jean-Pierre Mazat et Serge Nicolas, « Victor Henri: 111 years of his equation », Biochimie, vol. 107, , p. 161–166 (DOI 10.1016/j.biochi.2014.09.018, lire en ligne, consulté le )
- « Théorie générale de l'action de quelques diastases par Victor Henri [fac-similé publié dans C. R. Acad. Sci. Paris 135 (1902) 916–919] », Comptes Rendus Biologies, vol. 329, no 1, , p. 47–50 (DOI 10.1016/j.crvi.2005.10.007, lire en ligne, consulté le )
- Victor Henri, Lois générales de l'action des diastases (Thèse présentée à la faculté des sciences pour obtenir le grade de docteur Es-Sciences Naturelles), Paris, Librairie scientifique A Hermann,
- (en) Athel Cornish-Bowden, « One hundred years of Michaelis–Menten kinetics », Perspectives in Science, vol. 4, , p. 3–9 (DOI 10.1016/j.pisc.2014.12.002, lire en ligne, consulté le )
- (en) Kenneth A. Johnson et Roger S. Goody, « The Original Michaelis Constant: Translation of the 1913 Michaelis–Menten Paper », Biochemistry, vol. 50, no 39, , p. 8264–8269 (ISSN 0006-2960, DOI 10.1021/bi201284u, lire en ligne, consulté le )
- (en) Athel Cornish-Bowden, « The origins of enzyme kinetics », FEBS Letters, vol. 587, no 17, , p. 2725–2730 (ISSN 1873-3468, DOI 10.1016/j.febslet.2013.06.009, lire en ligne, consulté le )
- (en) George Edward Briggs et John Burdon Sanderson Haldane, « A Note on the Kinetics of Enzyme Action », Biochemical Journal, vol. 19, no 2, , p. 338–339 (ISSN 0264-6021 et 1470-8728, PMID 16743508, DOI 10.1042/bj0190338, lire en ligne, consulté le )
- (en) Donald D. Van Slyke et Glenn E. Cullen, « The mode of action of urease and of enzymes in general », J. Biol. Chem, vol. 19(2), , p. 141-180 (lire en ligne)
 Portail de la biochimie
Portail de la biochimie