Potentiel électrochimique de membrane
Toute cellule biologique est entourée d'une membrane dite membrane plasmique. Cette membrane est relativement imperméable aux espèces électriquement chargées telles que les ions et aux molécules qui peuvent participer à l'activité électrochimique (molécules polaires) telles que l'eau. Elle présente ainsi une grande résistance électrique et forme en quelque sorte un dipôle (comme un condensateur).
Grâce à ces propriétés, la membrane sépare en deux compartiments étanches l'intérieur de la cellule, le cytoplasme, de l'extérieur de la cellule, le milieu extracellulaire. De par l'activité permanente des protéines membranaires, la composition ionique de ces deux compartiments est différente. C'est pourquoi il existe, pour chacun des composés ioniques dont la concentration est différente, un potentiel électrochimique.
Équation de Nernst : Potentiel électrochimique pour chaque composé ionique
Soient deux compartiments de même volume contenant de l'eau pure et séparés par une membrane poreuse. Dans le compartiment A, on dissout une quantité de sel de cuisine (NaCl) et on laisse le compartiment B inchangé. Le sel se dissocie entièrement en Na+ et Cl- dans le compartiment A. La première loi de diffusion de Fick prédit qu'un flux de Na+ et Cl- va avoir lieu depuis le compartiment A vers le compartiment B jusqu’à équilibration des concentrations en Na+ et Cl- dans les deux compartiments. Afin de fixer les idées, si on introduit 1 mole de NaCl dans le compartiment A, à l'équilibre, il y aura 0.5 mole de NaCl dans chaque compartiment. Il s'agit du principe fondamental de la dialyse effectuée sur les insuffisants rénaux.
Supposons maintenant que la membrane ait des pores qui ne laissent passer que les ions Na+. On réitere l'expérience. On constate alors que la concentration en sel du compartiment B reste nulle. Si on place une électrode de mesure dans chaque compartiment, on remarque qu'une différence de potentiel entre le compartiment A et le compartiment B s'est établie. L'explication est la suivante : Les ions Na+ diffusent de A vers B. Au niveau de la membrane, les ions Na+ traversent, mais pas les ions Cl-. Une accumulation de charge négatives (Cl-) se fera du côté A et une accumulation de charges positives (Na+) se fera du côté B. Le champ électrique créé par cette séparation de charge s'oppose à toute entrée supplémentaire d'ions Na+. En fait, la diffusion continue mais elle est compensée exactement par une migration imposée par le champ électrique, il s'agit d'un équilibre dynamique. La valeur de la différence de potentiel entre les compartiments A et B est donnée par l'équation de Nernst.

où :
- Ei est le potentiel électrique du compartiment i
- [I]i la concentration de l'espèce perméante I du compartiment i (dans notre exemple Na+)
- R la constante des gaz parfaits
- T la température en kelvin
- z la charge de l'ion perméant (avec le signe)
- F la constante de Faraday
- (F = 96 485 C.mol-1 = N e où N est le nombre d'Avogadro et e la charge élémentaire de l'électron)
- ln le logarithme népérien.
Pour l'équation de Nernst telle qu'utilisée en électrochimie, voir Équation de Nernst.
Équation de Goldman-Hodgkin-Katz : Potentiel électrochimique pour plusieurs composés ioniques localement
L'Équation de Goldman-Hodgkin-Katz en voltage donne la valeur du potentiel de membrane lorsque plus d'une conductance est active au sein de la membrane considérée. Il s'agit typiquement du cas du neurone lors du potentiel d'action, mais aussi le cas de la majorité des cellules lorsque l'hypothèse simplificatrice de la seule activité de canaux potassiums n'est pas posée. On remarque en effet que la valeur mesurée du potentiel de membrane diffère de la valeur théorique donné par l'équation de Nernst appliquée au potassium.
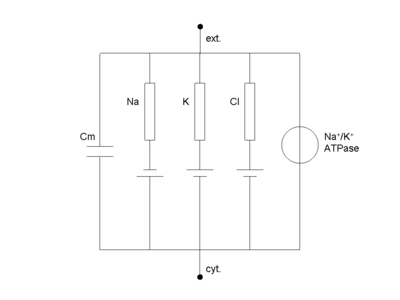
Équivalent électrique de la cellule
La cellule peut être décrite par un schéma électrique équivalent (voir figure ci-contre). On y distingue :
- Cm, la capacité de la membrane, égale à 1 µF.cm-2, directement proportionnelle à la surface de membrane
- les diverses conductances (rectangles) qui sont en général variables dans le temps
- les piles électrochimiques associées (donné par l'équation de Nernst)
- le générateur de tension (pompes membranaires, en particulier l'ubiquitaire pompe sodium-potassium)
Théories des câbles : propagation de potentiel le long de la membrane
L'axone du neurone est un prolongement cellulaire comportant des canaux ioniques en grande quantité. L'activité des canaux est responsable de la propagation de l'influx nerveux.
L'équivalent électrique de l'axone est une juxtaposition parallèle d'équivalents électriques reliés entre eux par une résistance longitudinale.
Propagation simple
Cette propagation se fait de proche en proche le long de l'axone, grâce à l'ouverture de canaux ioniques. Du fait de l'inhibition automatique d'un canal, il existe une période durant laquelle le canal est inutilisable pour propager le potentiel d'action, ce qui explique que ce potentiel d'action ne peut revenir en arrière. La vitesse de propagation est proportionnelle au diamètre du neurone sur lequel le potentiel d'action se propage. Ainsi il a été démontré que si les neurones oculaires ne possédaient pas de gaine de myéline, il faudrait que ces axones aient 30cm de diamètre pour propager un potentiel d'action à la même vitesse qu'in vivo.
Propagation saltatoire
La propagation saltatoire est due à l'existence, autour de certains axones, d'une gaine de myéline isolante. Les manchons de myéline sont séparés par des sections non myélinisées de l'axone : les nœuds de Ranvier. La propagation saltatoire permet une propagation beaucoup plus rapide des potentiels d'action, car ces derniers « sautent » d'un nœud de Ranvier à l'autre, d'où le nom de propagation « saltatoire ».
Rôles physiologiques
L'existence d'un potentiel électrochimique de part et d'autre de la membrane plasmique est universelle parmi les cellules. Il ne disparaît qu'à la mort de la cellule (Potentiel trans-membranaire = 0). Il est maintenu activement par des pompes ioniques, pompe sodium-potassium chez les cellules animales, pompes à protons chez les cellules végétales et les champignons. Il constitue une énergie potentielle qui est utilisée pour transporter plusieurs petites molécules contre leur gradient de concentration. Les nutriments (composés nécessaires au métabolisme cellulaire) sont importés, tandis que les déchets du métabolisme sont exportés.
Le potentiel électrochimique de membrane constitue aussi la force motrice nécessaire au courant potassique de fuite qui définit le potentiel de repos d'une cellule excitable, ainsi qu'aux courants sodiques, calciques et potassiques activés pendant un potentiel d'action ou un potentiel postsynaptique.
Au niveau de la sous-compartimentation de la cellule, d'autres potentiels électrochimiques sont maintenus en permanence :
- dans les organites producteurs d'énergie (mitochondries et chloroplastes), un gradient électrochimique de protons est maintenu de part et d'autre de la membrane interne, et est en équilibre avec la synthèse ou la dégradation d'ATP.
- de part et d'autre des membranes des réticulum endoplasmiques, en particulier les calciosomes remplis de calcium.
- dans certaines vésicules à faible pH.
Le potentiel diastolique maximal est la valeur la plus négative qui suit le potentiel d'action, sa valeur varie selon les cellules.
Différence de concentration ionique de part et d'autre de la membrane
| Ion | Concentration intracellulaire (mmol/l) | Concentration extracellulaire (mmol/l) | Rapport | Potentiel d'équilibre d'après l'équation de Nernst |
|---|---|---|---|---|
| Na+ | 20 | 140 | 1:7 | env. +51 mV |
| K+ | 150 | 4 | 38:1 | -97 mV |
| Ca2+ | 10-4 | 1.8 | +120 mV | |
| Cl- | 20 | 100 | -42 mV | |
| HCO3- | 8-10 | 26-28 | -27 mV | |
| Protéine anionique (chargée négativement) | 155 | 5 | -75 mV |
| Ion | Concentration intracellulaire (mmol/l) | Concentration extracellulaire (mmol/l) | Rapport | Potentiel d'équilibre d'après l'équation de Nernst |
|---|---|---|---|---|
| Na+ | 50 | 440 | 1:8 | +55 mV |
| K+ | 400 | 2 | 200:1 | -142 mV |
| Ca2+ | 10-7 | 14 | +149 mV | |
| Cl- | 52 | 560 | -60 mV | |
| HCO3- | ||||
| Protéine anionique (chargée négativement) | ||||
| Ion | Concentration intracellulaire (mmol/l) | Concentration extracellulaire (mmol/l) | Rapport | Potentiel d'équilibre d'après l'équation de Nernst |
|---|---|---|---|---|
| Na+ | 12 | 145 | 1:12 | +67 mV |
| K+ | 155 | 4 | 39:1 | -98 mV |
| Ca2+ | 10-7 | 1,5 | +129 mV | |
| Cl- | 4 | 123 | -90 mV | |
| HCO3- | ||||
| Protéine anionique (chargée négativement) | 155 | |||
Notes et références
- (de) Josef Dudel; Randolf Menzel; Robert F Schmidt, Randolf Menzel et Robert F Schmidt, Neurowissenschaft : vom Molekül zur Kognition, Berlin, Springer, , p.67
Articles connexes
- Équation de Nernst
- Potentiel de repos
- Potentiel d'action
- Potentiel postsynaptique
- Théorie chimiosmotique
- Gradient électrochimique

