Paléoneurologie
La paléoneurologie, ou neuropaléontologie, est l'étude de l'évolution du cerveau fondée sur l'analyse des moulages endocrâniens des vertébrés fossiles. Elle est considérée comme une sous-discipline de la paléontologie[1] et des neurosciences. Comme les autres branches de la paléontologie, elle donne un aperçu spécifique de l'évolution des espèces. La paléoneurologie suppose une connaissance du fonctionnement des cerveaux des êtres vivants actuels, sans laquelle il est impossible de faire des inférences sur le fonctionnement des cerveaux anciens[2].
Le cerveau étant formé de tissus mous qui se décomposent rapidement après la mort, il n'est généralement pas disponible. Toutefois, appuyant sur la surface interne du crâne, il y a laissé des empreintes qui subsistent parfois pendant plusieurs millions d'années. Les paléoneurologues analysent les moulages de l'intérieur du crâne, ou endocastes, qui reproduisent les détails de la morphologie externe des cerveaux imprimés sur les surfaces internes des crânes[3].
Les crânes fossiles et leurs moulages peuvent être comparés les uns aux autres ; ils peuvent être également confrontés aux crânes et fossiles d'individus modernes, et même comparés à ceux d'autres espèces pour faire des inférences sur l' anatomie fonctionnelle, la physiologie et la phylogénie.
Histoire

Anatomie comparée
Georges Cuvier, principal fondateur de l'anatomie comparée, qui se développe au XIXe siècle, peut être considéré comme un précurseur de la paléoneurologie[1]. Dans la cavité crânienne remplie de matrice sédimentaire d'un Anoplotherium (genre éteint de mammifère de la fin du Paléogène), il identifie un moulage endocrânien naturel[1]. Il décrit plusieurs moulages naturels récupérés dans les carrières à plâtre de la région parisienne. Il compare des moulages de cerveaux de vertébrés fossiles et ceux des vertébrés actuels. Il a souligné le fait que la fonction de l'organe doit coïncider avec sa forme[4].
Étude de l'évolution
L'anatomie comparée a été très influencée à la fin du XIXe siècle par les ouvrages de Charles Darwin, notamment par L'Origine des espèces paru en 1859. Ce travail modifie les vues des spécialistes d'anatomie comparée qui essayaient de comprendre la manière dont les différentes parties du cerveau avaient évolué[4].
Othniel Charles Marsh est un des premiers à tenter une étude systématique de l'évolution du cerveau des animaux fossiles[1]. Les « lois de Marsh » (1884) établissent ainsi que tous les mammifères de l'ère tertiaire avaient de petits cerveaux, qui ont connu au cours du temps un accroissement de taille graduel[1]. Les lois de Marsh ne sont plus considérées comme valides aujourd'hui mais elles ont marqué la paléoneurologie jusqu'au milieu du XXe siècleref name=Cours/>.
Usage du microscope
Une autre innovation majeure qui a contribué à la formation de la paléoneurologie est le microscope ; inventé au XVIIe siècle, il n'a été utilisé en biologie qu'à la fin du XIXe siècle. Les techniques d'observation des cellules cérébrales au microscope ont mis beaucoup de temps à s'affiner. En 1873, avec cet outil en main, Camillo Golgi commence à détailler le cerveau de manière cellulaire, mettant en évidence la structure du neurone ; il utilise des techniques de microscopie rendant accessible l'axone (prolongement du neurone qui conduit l'influx nerveux).
Neuranatomie comparée
Ludwig Edinger (1855-1918) établit une nouvelle branche de l'anatomie appelée neuroanatomie comparée. Il soutient que les vertébrés ont évolué dans une série progressive linéaire. Il pense également que les changements dans le cerveau sont basés sur une série d'ajouts et de différenciations et que les cerveaux les plus complexes sont ceux qui appartiennent aux espèces les plus encéphalisées (espèces qui ont un grand cerveau proportionnellement au poids total du corps)[5].
La période 1885-1935 est un moment de foisonnement d'idées en neuroanatomie comparée. Cette ère culmine avec la publication de "L'anatomie comparée du système nerveux" par Arienns, Kappers, Huber et Cosby, article UI a influencé la paléontologue Tilly Edinger, fille de Ludwig Edinger.
Tilly Edinger, fondatrice de la paléoneurologie
Ottilie "Tilly" Edinger (1897-1967) découvre au cours de ses études universitaires en zoologie, géologie et paléontologie, un moulage endocrânien naturel de Nothosaurus, un reptile marin de l'ère mésozoïque, sujet de son premier article en 1921, puis étudie le cerveau des chauves-souris fossiles (en 1926)[6]. Elle définit en 1929 le nouveau domaine de la paléoneurologie moderne dans Die fossilen Gehirne (Fossil Brains, Les cerveaux fossiles, titre qui renvoie aux moulages endocrâniens naturels)[6]. Son ouvrage sur l’évolution du cerveau des Equidés (Horse Brains, 1948) est considéré comme un travail important ; c'est un des premiers à retracer les transformations intervenues dans plusieurs zones cérébrales au sein d'une même lignée phylogénétique. Selon le paléoneurologue Ralph Holloway (en), « Tilly Edinger a découvert que le volume et l'organisation du cerveau étaient en quelque sorte liés » ; « il y a des périodes au cours desquelles le cerveau du cheval semble se réorganiser, et d'autres périodes au cours desquelles sa taille semble changer »[7].
Tilly Edinger a contribué à déterminer la mesure dans laquelle les moulages endocrâniens révèlent l'anatomie des cerveaux anciens ; ainsi l'endocaste crânien des crocodiles reflète de manière exacte les proportions des différentes régions du cerveau, mais ce n'est pas le cas de celui des tortues. Elle a aussi évalué la capacité des endocastes crâniens à renseigner sur les modes de vie des organismes éteints et sur l'accroissement de la taille du cerveau au cours du temps géologique — sujets qui sont encore aujourd'hui des thèmes de recherche importants[6].
T. Edinger travaille en Allemagne jusqu’en 1939 puis fuyant le nazisme elle émigre aux États-Unis où elle occupe un poste de chercheur au Harvard's Museum of Comparative Zoology[6].
Moulages endocrâniens
Les moulages endocrâniens révèlent les marques créées par la pression que le cerveau a exercée sur le crâne.
Moulages naturels

Les endocastes peuvent se former naturellement, lorsque des sédiments ont rempli la cavité crânienne au moment de l'enfouissement, et se sont durcis. Les endocastes naturels sont très rares ; le crâne de l'enfant de Taung et celui de Selam, l'enfant de Dikika[8], en sont chez les hominidés des exemples célèbres.
Moulages artificiels
Ils sont réalisés traditionnellement par injection d'un matériau de moulage dans la cavité crânienne. Ils peuvent être formés d'argile, de latex, de plâtre de Paris, de pâte à modeler ou de silicone. Aujourd'hui les moulages peuvent être effectués sur ordinateur[7] ; ces moulages numériques constituent une technique non invasive, particulièrement utile quand l'injection de matériau de moulage est impossible, parce que le crâne a été rempli de sédiments, et que cet endocaste naturel ne peut être retiré sans que les parties squelettiques du fossile ne soient détruites. L'imagerie par tomodensitométrie est obtenue grâce à l'application de rayons X pour produire des tomographies, ou des images en coupes, qui sont similaires aux images produites lors des IRM[9].
Les endocastes sont révélateurs mais ils ont leurs limites : ils présentent rarement des circonvolutions (reliefs) et les sillons (dépressions) en raison des membranes qui entouraient et protégeaient le tissu cérébral vivant (la pie-mère, la mère arachnoïde et la dure-mère). En outre, tous les endocastes ne sont pas créés à partir d'un fossile crânien complet et, de ce fait, les parties manquantes sont reconstituées de manière approximative sur la base de fossiles similaires. Dans certains cas, des fragments de plusieurs fossiles de la même espèce sont utilisés pour construire un seul endocaste[10].
Principaux sujets d'étude

Volume endocrânien
L'analyse statistique des moulages endocrâniens donne des informations sur les augmentations du volume cérébral global (le « volume endocrânien »).
L'évaluation de la capacité crânienne au fil du temps indique une augmentation du volume du cerveau depuis les premiers hominidés jusqu'à Homo sapiens ; le volume du cerveau des hommes modernes est ainsi trois à quatre fois supérieur à celui des australopithèques comme Lucy, fossile daté de 3,1 millions d'années[7]. Il y a 2 millions d'années, le cerveau des hominidés avait un volume de 600–750 cm3 ; celui d'Homo erectus, il y a environ 1,5 million d'années, est passé à 1000 cm3[7]. Aujourd'hui, le nôtre a en moyenne un volume de 1350 cm3 ; il aurait diminué d'environ 10% au cours des 30 000 dernières années ; notre cerveau est ainsi plus petit que celui de l'Homme de Néandertal[7].
La tendance générale à l'augmentation du volume endocrânien chez les hominidés n'est pourtant pas une loi absolue. Ainsi, il n'y pas de différence notable entre le volume endocrânien des australopithèques datés de 3 millions d'années, et celui de Homo floresiensis, daté de 18 000 ans[1]. De plus, d'après les moulages endocrâniens connus, le genre Paranthropus, regroupant des hominidés qui vivaient il y a 2,7 à 1 million d'années, aurait conservé au cours du temps un volume endocrânien stable[1]. Pour tenter d'expliquer la différence entre le genre Paranthropus et son contemporain le genre Homo, soumis aux mêmes conditions environnementales, mais dont la plupart des espèces ont connu une expansion du cerveau, certains scientifiques évoquent une différence de régime alimentaire : le régime à faible rendement énergétique de Paranthropus n'aurait pas permis l'augmentation de la taille de son encéphale, les intestins, organe énergivore, ayant accaparé une grande partie des ressources nutritives disponibles (c'est l'hypothèse de l’« expensive-tissue hypothesis » de Aiello et de Wheeler)[1].
Le volume du cerveau est important dans la littérature scientifique pour discuter de l'identification taxonomique, de la complexité comportementale, et de l'intelligence. Toutefois, le lien entre le volume endocrânien et les aspects comportementaux ou cognitifs demeure à beaucoup d'égards incertain. Chez l'homme moderne, la capacité crânienne peut varier jusqu'à 1000 cm3, sans aucune corrélation avec le comportement ou l'intelligence[11]. Ce degré de variation équivaut presque à l'augmentation totale du volume que révèle la comparaison des fossiles australopithèques et des humains modernes, et remet en question la validité du recours à la capacité crânienne comme mesure du développement des capacités cognitives[12].
Traditionnellement, les paléoneurologues se sont concentrés sur l'estimation du volume de l'ancien cerveau. Ces mesures leur ont permis de déterminer le rapport entre masse cérébrale et poids corporel (en) des différentes espèces[4].
Les paléoneurologues étudient le volume mais aussi l'organisation neurologique du cerveau, qui a évolué au cours du temps.
Asymétries
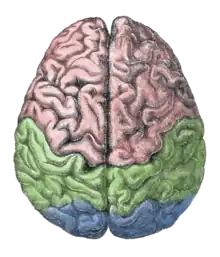
Lorsque le cerveau des hominidés a augmenté de volume il y a environ 2 millions d'années, des asymétries ont fait leur apparition dans les moulages endocrâniens. L'aire de Broca, une protubérance du côté gauche du lobe frontal associée à la production du langage articulé, apparaît également. L'asymétrie à elle seule ne suffit pas pour caractériser pas les humains ; on la trouve chez certains gorilles, et chez les oiseaux[13]. En revanche, la présence simultanée de plusieurs types d'asymétries serait propre aux homininés[14].
L'étude des asymétries entre les hémisphères droit et gauche intéresse donc tout particulièrement les paléoneurologues, car elles pourraient être liées à la préférence manuelle et au développement du langage. Les asymétries sont dues à la spécialisation des hémisphères cérébraux et sont observées à la fois de manière qualitative et quantitative. L'inégalité des hémisphères est appelée pétalia. Lorsque pour les lobes frontaux, un hémisphère se projette vers l'avant plus que l'autre, on parle de pétalia frontal ; le même phénomène peut se produire pour les lobes occipitaux, auquel cas on a affaire à un pétalia occipital[1]. L’hémisphère qui est plus projeté vers l'avant que l'autre a généralement aussi une extension latérale plus importante[14]. Les pétalias se produisent du fait de la tendance à utiliser la main droite ou la main gauche. Ils se produisent également en raison de la spécialisation de l'aire de Broca du lobe frontal chez les humains modernes[10]. Certaines asymétries ont été documentées sur des spécimens d' Homo erectus ; elles apparaissent déjà chez un spécimen d' Homo rudolfensis daté d'1,8 million d'années qui présente les mêmes asymétries que celles des humains modernes ; les scientifiques se posent la question de savoir si ces humains archaïques maîtrisaient le langage articulé[15].
Les paléoneurologues analysent les pétalias pour mesurer le degré de complexité de l'organisation du cerveau, mais ces asymétries ne constituent pas un indicateur définitif de l'évolution vers un cerveau plus humain.
Circonvolutions

Les circonvolutions céérébrales, les gyri (replis sinueux du cortex cérébral) et les sillons (dépressions de la surface du cortex) qui composent les plis du cerveau, sont dans un endocaste l'aspect le plus difficile à évaluer avec précision. La surface du cerveau apparaît souvent lisse et floue, à cause des méninges et du système vasculaire qui la recouvrent. Il est possible d'observer des modèles de gyri et de sillons (sulci) sous-jacents si un endocaste est correctement préservé, mais l'incertitude associée à ces modèles conduit souvent à la controverse[2]. Parce que les fossiles australopithèques robustes montrent ces détails, les circonvolutions sont incluses dans l'étude des endocastes chaque fois que cela est approprié[10].
Méninges
Bien que les méninges n'aient aucun lien avec le comportement, elles sont toujours étudiées dans le domaine de la paléoneurologie en raison du degré élevé de conservation des modèles méningés au sein d'une espèce qui peut servir de moyen de déterminer la taxonomie et la phylogénie[10].
Système vasculaire endocrânien
Parce que les vaisseaux sanguins méningés font partie de la couche la plus externe du cerveau, ils laissent souvent des sillons vasculaires dans la cavité crânienne qui sont capturés dans les endocastes. Le système vasculaire endocrânien permet de déduire la quantité de sang délivrée à différentes parties du cerveau[16].
Taille relative des lobes
Il est impossible de déterminer l'emplacement précis des sillons centraux ou précentraux à partir d'un endocaste. Cela peut néanmoins donner une idée approximative de la taille des lobes[4].
Limites
Le caractère lacunaire des archives fossiles empêche la paléoneurologie de documenter avec précision le cours de l'évolution du cerveau[17] De plus, la bonne préservation des fossiles est nécessaire pour garantir l'exactitude des endocastes étudiés[18], or les intempéries, l'érosion et la défiguration graduelle peuvent modifier les endocastes naturels comme les endocastes créés à partir de fossiles existants[19]. Autre limite : l'étude des endocastes s'arrête à l'anatomie externe des cerveaux exclusivement. Enfin, la paléoneurologie comparative ne révèle principalement que des différences de taille endocrânienne parmi les espèces apparentées. Puisqu'il n'y a pas de relation directe prouvée entre la taille du cerveau et l'intelligence, seules des inférences peuvent être faites concernant le développement du comportement d'anciens parents du genre Homo .
Ces limites de la paléoneurologie sont actuellement traitées par le développement d'outils plus avancés pour affiner l'étude des endocastes.
Nouvelles orientations
Maladies dégénératives et troubles fonctionnels
Selon le scientifique J. Ghika l'utilisation de la paléoneurologie est le meilleur moyen d'analyser plusieurs neurodégénérescences conduisant à des maladies telles que la maladie de Parkinson, les dyskinésies, les troubles de la marche, la maladie osseuse de Paget, la dystonie neurovégétative, etc. Une étude antérieure de SI Rapoport sur la maladie d'Alzheimer chez l'Homo sapiens a montré l'importance d'utiliser une perspective darwinienne pour mieux comprendre cette maladie elle-même et ses symptômes[20]. L'objectif est de déterminer les mécanismes génétiques qui conduisent à une atrophie cérébrale focale ou asymétrique affectant la démarche, les mouvements de la main (toute sorte de locomotion), le langage, la cognition, les troubles de l'humeur et du comportement.
Forme du cerveau, intelligence et performances cognitives
Des études récentes par Emiliano Bruner, Manuel Martin-Loechesb, Miguel Burgaletac et Roberto Colomc ont étudié le lien entre la forme du cerveau sagittal médian et la vitesse mentale. Selon les résultats de leur analyse, la corrélation est faible : l'influence de la géométrie du cerveau sagittal médian sur les performances cognitives individuelles est négligeable, mais fournit toujours des informations utiles sur les traits évolutifs du cerveau. Les zones associées au cortex pariétal semblent être impliquées dans les relations entre la géométrie du cerveau et la vitesse mentale[21].
Paléoneurologues célèbres
- Othniel Charles Marsh
- Tilly Edinger
- Harry Jerison, établit un quotient d'encéphalisation
- Leonard B. Radinsky (en), travaille sur l'endocrâne des mammifères
- James Hopson, travaille sur l'endocrâne des reptiles
- Peter Galton, sur l'endocrâne des dinosaures
- Emily Giffin-Buchholtz, sur les moulages endocrâniens des pachycéphalosaures
- Lawrence Witmer (en), endocrânes des dinosaures
- Dean Falk
- Ralph Holloway (en)
- Emiiano Bruner
- Antoine Balzeau
- Amélie Beaudet
- Pascaline Lauters
Voir également
Références
- Michel Brunet, « Paléontologie humaine », L’annuaire du Collège de France. Cours et travaux, no 111, , p. 349–374 (ISSN 0069-5580, DOI 10.4000/annuaire-cdf.1482, lire en ligne, consulté le )
- Bruner, « Fossil traces of the human thought: paleoneurobiology and the evolution of the genus Homo », Journal of Anthropologia Sciences, vol. 81, , p. 29–56 (lire en ligne[archive du ], consulté le )
- Falk, « Hominid Paleoneurobiology », Annual Review of Anthropology, vol. 16, , p. 13–30 (DOI 10.1146/annurev.an.16.100187.000305, JSTOR 2155862)
- Ralph L. Holloway, Chet C. Sherwood, Patrick R. Hof et James K. Rilling, Encyclopedia of Neuroscience, , 1326–1334 p. (ISBN 978-3-540-23735-8, DOI 10.1007/978-3-540-29678-2_3152), « Evolution of the Brain in Humans – Paleoneurology »
- Northcutt, « Changing Views of Brain Evolution », Brain Research Bulletin, vol. 55, no 6, , p. 663–674 (PMID 11595351, DOI 10.1016/S0361-9230(01)00560-3)
- Buchholtz et Seyfarth, « The Study of "Fossil Brains": Tilly Edinger (1897–1967) and the Beginnings of Paleoneurology », BioScience, vol. 51, no 8, , p. 674 (ISSN 0006-3568, DOI 10.1641/0006-3568(2001)051[0674:TSOFBT]2.0.CO;2)
- (en) « Traquer l'évolution du cerveau humain à travers les moulages du crâne intérieur »,
- Kate Wong, « Une fillette comme aïeule », sur Pourlascience.fr (consulté le )
- Marino, Uhen, Pyenson et Frohlich, « Reconstructing Cetacean Brain Evolution Using Computed Tomography », The Anatomical Record Part B: The New Anatomist, vol. 272B, no 1, , p. 107–17 (PMID 12731077, DOI 10.1002/ar.b.10018, lire en ligne)
- Ralph L. Holloway, Douglas C. Broadfield et Michael S. Yuan, The Human Fossil Record, Volume Three : Brain Endocasts--The Paleoneurological Evidence, Wiley-Liss, , 315 p. (ISBN 978-0-471-41823-8, lire en ligne)
- Chez Homo sapiens, «il n'existe aucun rapport entre le volume du cerveau et les capacités intellectuelles. Des exemples fameux sont les cerveaux d'Anatole France et d'Ivan Tourgueniev : le premier pesait 1kg et le second 2kg », « biosex: Sexe et cerveau », sur biosex.univ-paris1.fr (consulté le )
- Holloway, « Cranial Capacity, Neural Reorganization, and Hominid Evolution: A Search for More Suitable Parameters », American Anthropologist, vol. 68, no 1, , p. 103–21 (DOI 10.1525/aa.1966.68.1.02a00090)
- Habib Michel, « Développement de la dominance cérébrale : revue des données disponibles et proposition d'une hypothèse originale », Développements, 2009/2 (n° 2), p. 5-26. DOI : 10.3917/devel.002.0005. URL : https://www.cairn.info/revue-developpements-2009-2-page-5.htm
- (en) Antoine Balzeau et Emmanuel Gilissen, « Endocranial shape asymmetries in Pan paniscus, Pan troglodytes and Gorilla gorilla assessed via skull based landmark analysis », Journal of Human Evolution, vol. 59, no 1, , p. 54–69 (ISSN 0047-2484, DOI 10.1016/j.jhevol.2010.03.013, lire en ligne, consulté le )
- « Modern humans also show asymmetries in the Broca’s cap regions of the third inferior convolution of the frontal cortex. These asymmetries probably differ by handedness as well as by unknown functional relationships. Such asymmetries are present in Neandertals and even earlier on some Homo erectus specimens (indeed they are clear on the 1.8 million year-old Homo rudolfensis specimen, KNM-ER 1470). They cannot prove that this or that hominid had language, but if these asymmetries are homologous to those found in modern humans, well, why not? », Ralph L. Holloway, Chet C. Sherwood, Patrick R. Hof et James K. Rilling, Encyclopedia of Neuroscience, , 1326–1334 p. (ISBN 978-3-540-23735-8, DOI 10.1007/978-3-540-29678-2_3152), « Evolution of the Brain in Humans – Paleoneurology »
- Dominique Grimaud-Hervé, The Human Fossil Record, Volume Three : Brain Endocasts--The Paleoneurological Evidence, Wiley-Liss, , 315 p. (ISBN 978-0-471-41823-8, lire en ligne), « Part Five - Endocranial Vasculature »
- Rogers, « Reconstructing the Behaviors of Extinct Species: An Excursion Into Comparative Paleoneurology », American Journal of Medical Genetics, vol. 134A, no 4, , p. 349–56 (PMID 15759265, DOI 10.1002/ajmg.a.30538)
- Emiliano Bruner, Giorgio Manzi et Juan Luis Arsuaga, Encephalization and allometric trajectories in the genus Homo : Evidence from the Neandertal and modern lineages, vol. 100 (no 26), , 15335–15340 p. (ISBN 978-2-536-67110-7, PMID 14673084, DOI 10.1073/pnas.2536671100, Bibcode 2003PNAS..10015335B, lire en ligne)
- Bruner, « Geometric morphometrics and paleoneurology: brain shape evolution in the genus Homo », Journal of Human Evolution, vol. 47, no 5, , p. 279–303 (PMID 15530349, DOI 10.1016/j.jhevol.2004.03.009, lire en ligne[archive du ], consulté le )
- Rapoport, « Brain evolution and Alzheimer's disease », Review Neurology, paris, vol. 144, no 2, , p. 79–90 (PMID 2898165)
- Bruner, Manuel Martin-Loeches, Miguel Burgaleta et Roberto Colom, « Midsagittal brain shape correlation with intelligence and cognitive performance », Intelligence, vol. 39, nos 2–3, march–april 2011, p. 141–147 (DOI 10.1016/j.intell.2011.02.004, lire en ligne)
Bibliographie
- (en) Emiliano Bruner, Human Paleoneurology, Cham, Springer, (ISBN 978-3-319-08500-5, lire en ligne)
- (en) Ralph L. Holloway, Chet C. Sherwood, Patrick R. Hof et James K. Rilling, Encyclopedia of Neuroscience, , 1326–1334 p. (ISBN 978-3-540-23735-8, DOI 10.1007/978-3-540-29678-2_3152), « Evolution of the Brain in Humans – Paleoneurology »
- Antoine Balzeau, « Première description de l’endocrâne de Cro-Magnon 1 et étude de la variation et de l’évolution du cerveau chez les Hommes anatomiquement modernes », Bull. Mém. Soc. Anthropol. Paris, (lire en ligne)
- (en) Fabien Knoll et Soichiro Kawabe, « Avian palaeoneurology: Reflections on the eve of its 200th anniversary », Journal of Anatomy, vol. 236, no 6, , p. 965–979 (ISSN 1469-7580, PMID 31999834, PMCID PMC7219626, DOI 10.1111/joa.13160, lire en ligne, consulté le )
- (en) Emily A. Buchholtz et Ernst-August Seyfarth, « The Study of "Fossil Brains": Tilly Edinger (1897–1967) and the Beginnings of Paleoneurology », BioScience, vol. 51, no 8, , p. 674 (ISSN 0006-3568, DOI 10.1641/0006-3568(2001)051[0674:TSOFBT]2.0.CO;2)
 Portail de la paléontologie
Portail de la paléontologie  Portail de la Préhistoire
Portail de la Préhistoire  Portail des neurosciences
Portail des neurosciences  Portail origine et évolution du vivant
Portail origine et évolution du vivant