Enjambement (génétique)
L’enjambement, appelé également « entrecroisement » ou encore par les anglicismes « crossover » ou « crossing-over », est un phénomène génétique qui a lieu lors de la méiose et qui contribue au brassage génétique lors de la reproduction (Recombinaison génétique)[1].
Pour les articles homonymes, voir Crossover et Enjambement.


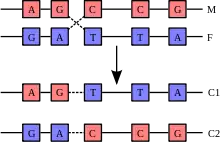
À la fin de la prophase 1 (début métaphase 1) de la méiose, les chromosomes homologues (i.e. d'une même paire) se chevauchent formant ainsi des figures caractéristiques appelées « tétrade de chromatides » et échangent des fragments de chromatides. Le résultat en est le brassage intra-chromosomique des allèles : tous les gènes situés sur une paire de chromosomes peuvent être brassés grâce au phénomène de la « recombinaison homologue par enjambement », ce qui modifie l'association d'allèles portée par chacun des chromosomes. Ainsi, les deux chromosomes de chaque paire après la méiose sont différents des deux chromosomes de départ, on dit qu'ils sont « recombinés ». Ce processus s’enclenche au cours de l'appariement et de la formation complexe synaptonémal et se termine pendant que les chromosomes homologues sont en synapsis.
Pour que l'échange ait lieu, les deux chromosomes de chaque paire se rapprochent et se brisent au même niveau de la séquence génétique pour ensuite échanger les fragments résultants. Ceci se fait au niveau des chiasmas, points de chevauchement de deux chromatides non-sœurs, observables en microscopie.
Il arrive parfois que les deux chromosomes ne se brisent pas au même niveau, ce qui entraîne une duplication de gènes sur un chromosome et une suppression sur l'autre. On appelle ce phénomène un enjambement inégal. Si un chromosome se brise de part et d'autre du même centromère et que les deux parties se rejoignent en excluant le centromère, un chromosome peut être perdu durant la division cellulaire.
Chaque paire de chromosomes réalise plusieurs enjambements durant la méiose, plus ou moins nombreux selon l'espèce et la longueur des chromosomes. La fréquence des enjambements dépend de la distance entre les gènes. Ainsi, plus des gènes sont distants du centromère et plus la probabilité pour qu'un crossing over survienne entre les 2 loci (=emplacement du gène sur le chromosome) est élevée.
L'enjambement a été découvert en 1909 par le Professeur belge Frans Alfons Janssens de l'Université catholique de Louvain qui l'avait appelé chiasmatypie et a été élaboré en théorie par Thomas Hunt Morgan qui publia ses observations en 1916 dans son ouvrage A Critique of the Theory of Evolution (Une Critique de la Théorie de l'Évolution), et la base physique de ce phénomène fut démontrée en 1931 par Harriet Creighton et Barbara McClintock[2].
Morgan est l'inventeur d'une unité de mesure (le centimorgan, symbole cM) de la distance entre deux gènes sur un même chromosome. Cette distance est calculée en fonction de la fréquence des enjambements ayant lieu entre ces deux gènes : 1 centimorgan correspond à 1 % de descendants au génotype recombiné (résultant d'un enjambement).
Notes et références
- [flash] Animation de biologie montrant le déroulement de l'enjambement au cours de la méiose.
- (en) Creighton H, McClintock B, « "A Correlation of Cytological and Genetical Crossing-Over in Zea Mays" », Proc Natl Acad Sci USA, no 17 (8), , p. 492–7 (lire en ligne)
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire